Provides interaction between the cell and the environment. Relationship between the body and the environment
The connection of the organism with the environment, from a physicochemical point of view, is an open system, that is, a system where biochemical processes are ongoing. The starting substances come from the environment, and the substances that are also continuously formed are carried outside. The equilibrium between the speed and concentration of products of multidirectional reactions in the body is conditional, imaginary, since the intake and removal of substances does not stop. Continuous connection with the environment allows us to consider a living organism as an open system.
For all living cells, the source of energy is the Sun. Plant cells capture energy from sunlight with the help of chlorophyll, using it for assimilation reactions during the process of photosynthesis. Cells of animals, fungi, and bacteria use solar energy indirectly, during the breakdown of organic substances synthesized by earthly plants.
Some of the cell's nutrients are broken down during cellular respiration, thus supplying the energy necessary for various types of cellular activity. This process takes place in organelles called mitochondria. Mitochondria consists of two membranes: the outer one, separating the organelle from the cytoplasm, and the inner one, forming numerous folds. The main product of respiration is ATP. It leaves the mitochondria and is used as an energy source for many chemical reactions in the cytoplasm and cell membrane. If oxygen is required for cellular respiration, then respiration is called aerobic, but if reactions occur in the absence of oxygen, then we speak of anaerobic respiration.
For any type of work performed in a cell, energy is used in one and only form - in the form of energy from the phosphate bonds of ATP. ATP is an easily mobile compound. The formation of ATP occurs on the inner membrane of mitochondria. ATP is synthesized in all cells during respiration due to the energy of oxidation of carbohydrates, fats and other organic substances. In green plant cells, the main amount of ATP is synthesized in chloroplasts due to solar energy. During photosynthesis, they produce many times more ATP than mitochondria. ATP decomposes with the rupture of phosphorus-oxygen bonds and the release of energy. This occurs under the action of the enzyme ATPase during the hydrolysis of ATP - the addition of water with the elimination of a phosphoric acid molecule. As a result, ATP is converted into ADP, and if two molecules of phosphoric acid are split off, then into AMP. The reaction of elimination of each gram-molecule of acid is accompanied by the release of 40 kJ. This is a very large energy output, which is why the phosphorus-oxygen bonds of ATP are usually called macroergistic (high-energy).
The use of ATP in plastic exchange reactions is carried out by coupling them with ATP hydrolysis. Molecules of various substances are charged with energy by attaching the phosphorus group released during hydrolysis from the ATP molecule, i.e. by phosphorylation.
The peculiarity of phosphate derivatives is that they cannot leave the cell, although their “discharged” forms freely pass through the membrane. Thanks to this, phosphorylated molecules remain in the cell until they are used in appropriate reactions.
The reverse process of converting ADP into ATP occurs by adding a phosphoric acid molecule to ADP, releasing water and absorbing a large amount of energy.
Thus, ATP is a universal and direct source of energy for cell activity. This creates a single cellular pool of energy and makes it possible to redistribute and transport it from one area of the cell to another.
The transfer of the phosphate group plays an important role in chemical reactions such as the assembly of macromolecules from monomers. For example, amino acids can be combined into peptides only after being previously phosphorylated. Mechanical processes of contraction or movement, transport of a dissolved substance against a concentration gradient and other processes involve the consumption of energy stored in ATP.
The process of energy metabolism can be represented as follows. High-molecular organic substances in the cytoplasm are enzymatically, by hydrolysis, converted into simpler ones from which they consist: proteins - into amino acids, poly- and disaccharides - into monosaccharides (+ glucose), fats into glycerol and fatty acids. There are no oxidative processes, little energy is released, which is not used and goes into thermal form. Most cells use carbohydrates first. Polysaccharides (starch in plants and glycogen in animals) are hydrolyzed to glucose. Glucose oxidation occurs in three phases: glycolysis, oxidative decarboxylation (Krebs cycle - citric acid cycle) and oxidative phosphorylation (respiratory chain). Glycolysis, as a result of which one molecule of glucose is split into two molecules of pyruvic acid with the release of two molecules of ATP, occurs in the cytoplasm. In the absence of oxygen, pyruvic acid is converted to either ethanol (fermentation) or lactic acid (anaerobic respiration).
When glycolysis occurs in animal cells, the six-carbon molecule of glucose breaks down into two molecules of lactic acid. This process is multi-stage. It is carried out sequentially by 13 enzymes. During alcoholic fermentation, two molecules of ethanol and two molecules of CO2 are formed from a glucose molecule.
Glycolysis is a phase common to anaerobic and aerobic respiration; the other two occur only under aerobic conditions. The process of oxygen-free oxidation, in which only part of the energy of metabolites is released and used, is final for anaerobic organisms. In the presence of oxygen, pyruvic acid passes into the mitochondria, where, as a result of a number of sequential reactions, it is completely oxidized aerobically to H2O and CO2 with simultaneous phosphorylation of ADP to ATP. In this case, two ATP molecules are produced by glycolysis, two by the Krebs cycle, and 34 by the respiratory chain. The net yield for the complete oxidation of one glucose molecule to H2O and CO2 is 38 molecules.
Thus, in aerobic organisms, the final decomposition of organic substances is carried out by oxidizing them with atmospheric oxygen to simple inorganic substances: CO2 and H2O. This process takes place on the cristae of mitochondria. In this case, the maximum amount of free energy is released, a significant part of which is reserved in ATP molecules. It is easy to see that aerobic oxidation provides the cell with free energy to the greatest extent.
As a result of catabolism, energy-rich ATP molecules accumulate in the cell, and CO2 and excess water are released into the external environment.
Sugar molecules not required for respiration can be stored in the cell. Excess lipids are either broken down, after which the products of their breakdown enter the mitochondria as a substrate for respiration, or are deposited as reserves in the cytoplasm in the form of fat droplets. Proteins are built from amino acids entering the cell. Protein synthesis occurs in organelles called ribosomes. Each ribosome consists of two subparticles - large and small: both subparticles include protein molecules and RNA molecules.
Ribosomes are often attached to a special membrane system consisting of cisterns and vesicles - the so-called endoplasmic reticulum (ER); in cells that produce a lot of protein, the endoplasmic reticulum is often very well developed and covered with ribosomes. Some enzymes are only effective if they are attached to a membrane. Most of the enzymes involved in lipid synthesis are located here. Thus, the endoplasmic reticulum is like a kind of cell workbench.
In addition, the ER divides the cytoplasm into separate compartments, i.e., it separates various chemical processes occurring simultaneously in the cytoplasm, and thereby reduces the likelihood that these processes will interfere with each other.
Products produced by a given cell are often used outside the cell. In such cases, proteins synthesized on ribosomes pass through the membranes of the endoplasmic reticulum and are packaged into membrane vesicles that form around them, which are then detached from the ER. These vesicles, flattened and stacked on top of each other, like stacked pancakes, form a characteristic structure called the Golgi complex, or Golgi apparatus. During their stay in the Golgi apparatus, proteins undergo certain changes. When the time comes for them to leave the cell, the membrane vesicles merge with the cell membrane and are emptied, pouring their contents out, i.e., secretion occurs by exocytosis.
The Golgi apparatus also produces lysosomes - membrane sacs containing digestive enzymes. Finding out how a cell makes, packages, and exports certain proteins, and how it “knows” which proteins it should keep for itself, is one of the most fascinating branches of modern cytology.
The membranes of any cell are constantly moving and changing. ER membranes move slowly throughout the cell. Individual sections of these membranes separate and form vesicles, which temporarily become part of the Golgi apparatus, and then, through the process of exocytosis, merge with the cell membrane.
Later, the membrane material is returned to the cytoplasm, where it is used again.
The exchange of substances entering the cell or released by it outside, as well as the exchange of various signals with the micro- and macroenvironment, occurs through the outer membrane of the cell. As is known, the cell membrane is a lipid bilayer into which various protein molecules are embedded that act as specialized receptors, ion channels, devices that actively transport or remove various chemicals, intercellular contacts, etc. In healthy eukaryotic cells, phospholipids are distributed in the membrane asymmetrically: the outer surface consists of sphingomyelin and phosphatidylcholine, the inner surface - of phosphatidylserine and phosphatidylethanolamine. Maintaining such asymmetry requires energy expenditure. Therefore, in the event of cell damage, infection, or energy starvation, the outer surface of the membrane is enriched with phospholipids that are unusual for it, which becomes a signal for other cells and enzymes about cell damage with a corresponding reaction to this. The most important role is played by the soluble form of phospholipase A2, which breaks down arachidonic acid and creates lysoforms from the above-mentioned phospholipids. Arachidonic acid is the limiting link for the creation of inflammatory mediators such as eicosanoids, and protective molecules - pentraxins (C-reactive protein (CRP), precursors of amyloid proteins) - are attached to lysoforms in the membrane, followed by activation of the complement system along the classical pathway and cell destruction.
The structure of the membrane helps preserve the characteristics of the internal environment of the cell, its differences from the external environment. This is ensured by the selective permeability of the cell membrane and the existence of active transport mechanisms in it. Their disruption as a result of direct damage, for example, by tetrodotoxin, ouabain, tetraethylammonium, or in the case of insufficient energy supply to the corresponding “pumps” leads to disruption of the electrolyte composition of the cell, changes in its metabolism, disruption of specific functions - contraction, conduction of excitation impulses, etc. Disturbance of cellular ion channels (calcium, sodium, potassium and chloride) in humans can also be genetically determined by mutation of the genes responsible for the structure of these channels. So-called channelopathies cause hereditary diseases of the nervous, muscular, and digestive systems. Excessive entry of water into the cell can lead to its rupture - cytolysis - due to perforation of the membrane when complement is activated or an attack by cytotoxic lymphocytes and natural killer cells.
The cell membrane has many receptors built into it - structures that, when combined with the corresponding specific signaling molecules (ligands), transmit a signal inside the cell. This occurs through various regulatory cascades consisting of enzymatically active molecules that are sequentially activated and ultimately contribute to the implementation of various cellular programs, such as growth and proliferation, differentiation, motility, aging, and cell death. Regulatory cascades are quite numerous, but their number has not yet been fully determined. The system of receptors and regulatory cascades associated with them also exists inside the cell; they create a specific regulatory network with points of concentration, distribution and selection of the further signal path depending on the functional state of the cell, the stage of its development, and the simultaneous action of signals from other receptors. The result of this may be inhibition or strengthening of the signal, directing it along a different regulatory pathway. Both the receptor apparatus and signal transduction pathways through regulatory cascades, for example to the nucleus, can be disrupted as a result of a genetic defect that occurs as a congenital defect at the organismal level or due to a somatic mutation in a specific cell type. These mechanisms can be damaged by infectious agents, toxins, and also change during the aging process. The final stage of this may be a disruption of the functions of the cell, the processes of its proliferation and differentiation.
On the surface of cells there are also molecules that play an important role in the processes of intercellular interaction. These may include cell adhesion proteins, histocompatibility antigens, tissue-specific, differentiating antigens, etc. Changes in the composition of these molecules cause disruption of intercellular interactions and can cause the activation of appropriate mechanisms for the elimination of such cells, because they pose a certain danger to the integrity of the body as reservoir of infection, especially viral, or as potential initiators of tumor growth.
Violation of the energy supply of the cell
The source of energy in the cell is food, after the breakdown of which energy is released into final substances. The main place of energy production is mitochondria, in which substances are oxidized with the help of enzymes of the respiratory chain. Oxidation is the main supplier of energy, since as a result of glycolysis, no more than 5% of energy is released from the same amount of oxidation substrates (glucose), compared to oxidation. About 60% of the energy released during oxidation is accumulated by oxidative phosphorylation in high-energy phosphates (ATP, creatine phosphate), the rest is dissipated as heat. Subsequently, high-energy phosphates are used by the cell for processes such as pump operation, synthesis, division, movement, secretion, etc. There are three mechanisms, damage to which can cause a disruption in the cell’s energy supply: the first is the mechanism of synthesis of energy metabolism enzymes, the second is the mechanism of oxidative phosphorylation , the third is the mechanism of energy use.
Disruption of electron transport in the mitochondrial respiratory chain or uncoupling of ADP oxidation and phosphorylation with loss of proton potential, the driving force for ATP generation, leads to a weakening of oxidative phosphorylation in such a way that most of the energy is dissipated as heat and the number of high-energy compounds decreases. The uncoupling of oxidation and phosphorylation under the influence of adrenaline is used by the cells of homeothermic organisms to increase heat production while maintaining a constant body temperature during cooling or increasing it during fever. Significant changes in mitochondrial structure and energy metabolism are observed in thyrotoxicosis. These changes are initially reversible, but after a certain point they become irreversible: mitochondria fragment, disintegrate or swell, lose cristae, turning into vacuoles, and eventually accumulate substances such as hyaline, ferritin, calcium, lipofuscin. In patients with scurvy, mitochondria fuse to form chondriospheres, possibly due to membrane damage by peroxide compounds. Significant damage to mitochondria occurs under the influence of ionizing radiation during the transformation of a normal cell into a malignant one.
Mitochondria are a powerful depot of calcium ions, where its concentration is several orders of magnitude higher than that in the cytoplasm. When mitochondria are damaged, calcium enters the cytoplasm, causing activation of proteinases with damage to intracellular structures and disruption of the functions of the corresponding cell, for example, calcium contractures or even “calcium death” in neurons. As a result of disruption of the functional capacity of mitochondria, the formation of free radical peroxide compounds sharply increases, which have a very high reactivity and therefore damage important components of the cell - nucleic acids, proteins and lipids. This phenomenon is observed during so-called oxidative stress and can have negative consequences for the existence of the cell. Thus, damage to the outer membrane of the mitochondria is accompanied by the release into the cytoplasm of substances contained in the intermembrane space, primarily cytochrome C and some other biologically active substances, which trigger chain reactions that cause programmed cell death - apoptosis. By damaging mitochondrial DNA, free radical reactions distort the genetic information necessary for the formation of certain respiratory chain enzymes, which are produced specifically in mitochondria. This leads to even greater disruption of oxidative processes. In general, the mitochondria's own genetic apparatus, compared to the genetic apparatus of the nucleus, is less protected from harmful influences that can change the genetic information encoded in it. As a result, dysfunction of mitochondria occurs throughout life, for example, during the aging process, during malignant transformation of the cell, as well as against the background of hereditary mitochondrial diseases associated with mutation of mitochondrial DNA in the egg. Currently, over 50 mitochondrial mutations have been described that cause hereditary degenerative diseases of the nervous and muscular systems. They are transmitted to the child exclusively from the mother, since the mitochondria of the sperm are not part of the zygote and, accordingly, the new organism.
Violation of the preservation and transmission of genetic information
The cell nucleus contains most of the genetic information and thereby ensures its normal functioning. Through selective gene expression, it coordinates cell activity during interphase, stores genetic information, and recreates and transmits genetic material during cell division. DNA replication and RNA transcription occur in the nucleus. Various pathogenic factors, such as ultraviolet and ionizing radiation, free radical oxidation, chemicals, viruses, can damage DNA. It is estimated that each cell of a warm-blooded animal takes 1 day. loses more than 10,000 bases. Here we should add violations when copying during division. If these damages persisted, the cell would not be able to survive. Protection lies in the existence of powerful repair systems, such as ultraviolet endonuclease, repair replication and recombination repair systems, which replace DNA damage. Genetic defects in repair systems cause the development of diseases caused by increased sensitivity to factors that damage DNA. This is xeroderma pigmentosum, as well as some accelerated aging syndromes, accompanied by an increased tendency to develop malignant tumors.
The system for regulating the processes of DNA replication, transcription of messenger RNA (mRNA), and translation of genetic information from nucleic acids into the structure of proteins is quite complex and multi-level. In addition to the regulatory cascades that trigger the action of transcription factors with a total number of over 3000, which activate certain genes, there is also a multi-level regulatory system mediated by small RNA molecules (interfering RNA; RNAi). The human genome, which consists of approximately 3 billion purine and pyrimidine bases, contains only 2% of the structural genes responsible for protein synthesis. The rest provide the synthesis of regulatory RNAs, which, simultaneously with transcription factors, activate or block the work of structural genes at the DNA level in chromosomes or influence the processes of translation of messenger RNA (mRNA) during the formation of a polypeptide molecule in the cytoplasm. Violation of genetic information can occur both at the level of structural genes and the regulatory part of DNA with corresponding manifestations in the form of various hereditary diseases.
Recently, much attention has been attracted to changes in genetic material that occur during the individual development of an organism and are associated with inhibition or activation of certain sections of DNA and chromosomes due to their methylation, acetylation and phosphorylation. These changes persist for a long time, sometimes throughout the entire life of the organism from embryogenesis to old age, and are called epigenomic heredity.
The proliferation of cells with altered genetic information is also prevented by systems (factors) that control the mitotic cycle. They interact with cyclin-dependent protein kinases and their catalytic subunits - cyclins - and block the cell from going through the full mitotic cycle, stopping division at the border between the presynthetic and synthetic phases (G1/S block) until DNA repair is completed, and if this is impossible, they initiate programmed death cells. These factors include the p53 gene, the mutation of which causes loss of control over the proliferation of transformed cells; it is observed in almost 50% of human cancers. The second checkpoint of the mitotic cycle is at the G2/M border. Here, the correct distribution of chromosomal material between daughter cells in mitosis or meiosis is controlled using a set of mechanisms that control the cell spindle, center and centromeres (kinetochores). The ineffectiveness of these mechanisms leads to disruption of the distribution of chromosomes or their parts, which is manifested by the absence of any chromosome in one of the daughter cells (aneuploidy), the presence of an extra chromosome (polyploidy), the separation of a part of a chromosome (deletion) and its transfer to another chromosome (translocation) . Such processes are very often observed during the proliferation of malignantly degenerated and transformed cells. If this happens during meiosis with germ cells, it leads either to the death of the fetus at an early stage of embryonic development, or to the birth of an organism with a chromosomal disease.
Uncontrolled cell proliferation during tumor growth occurs as a result of mutations in genes that control cell proliferation and are called oncogenes. Among more than 70 currently known oncogenes, most belong to components of cell growth regulation, some are represented by transcription factors that regulate gene activity, as well as factors that inhibit cell division and growth. Another factor limiting the excessive expansion (spread) of proliferating cells is the shortening of the ends of chromosomes - telomeres, which are not able to fully replicate as a result of purely steric interaction, therefore, after each cell division, the telomeres are shortened by a certain part of the bases. Thus, proliferating cells of an adult organism after a certain number of divisions (usually from 20 to 100 depending on the type of organism and its age) exhaust the telomere length and further chromosome replication stops. This phenomenon does not occur in sperm epithelium, enterocytes and embryonic cells due to the presence of the enzyme telomerase, which restores telomere length after each division. In most cells of adult organisms, telomerase is blocked, but, unfortunately, it is activated in tumor cells.
The connection between the nucleus and the cytoplasm and the transport of substances in both directions are carried out through pores in the nuclear membrane with the participation of special transport systems that consume energy. In this way, energy and plastic substances, signaling molecules (transcription factors) are transported to the nucleus. The reverse flow carries into the cytoplasm molecules of mRNA and transfer RNA (tRNA), ribosomes necessary for protein synthesis in the cell. The same route of transport of substances is also inherent in viruses, in particular such as HIV. They transfer their genetic material into the nucleus of the host cell with its further incorporation into the host genome and the transfer of newly formed viral RNA into the cytoplasm for further synthesis of proteins of new viral particles.
Violation of synthesis processes
Protein synthesis processes occur in cisterns of the endoplasmic reticulum, closely connected with pores in the nuclear membrane, through which ribosomes, tRNA and mRNA enter the endoplasmic reticulum. Here, the synthesis of polypeptide chains is carried out, which subsequently acquire their final form in the agranular endoplasmic reticulum and the lamellar complex (Golgi complex), where they undergo post-translational modification and combine with carbohydrate and lipid molecules. Newly formed protein molecules do not remain at the site of synthesis, but through a complex regulated process called proteinkinesis, are actively transferred to that isolated part of the cell where they will perform their intended function. In this case, a very important step is the structuring of the transferred molecule into an appropriate spatial configuration capable of performing its inherent function. This structuring occurs with the help of special enzymes or on a matrix of specialized protein molecules - chaperones, which help the protein molecule, newly formed or changed due to external influence, to acquire the correct three-dimensional structure. In the event of an adverse effect on the cell, when there is a possibility of disruption of the structure of protein molecules (for example, with an increase in body temperature, an infectious process, intoxication), the concentration of chaperones in the cell increases sharply. Therefore, such molecules are also called stress proteins, or heat shock proteins. Violation of the structuring of a protein molecule leads to the formation of chemically inert conglomerates, which are deposited in the cell or outside it during amyloidosis, Alzheimer's disease, etc. Sometimes a pre-structured similar molecule can serve as a matrix, and in this case, if the primary structuring occurs incorrectly, all subsequent molecules also will be defective. This situation occurs in so-called prion diseases (scrapie in sheep, rabid cows, kuru, Creutzfeldt-Jakob disease in humans), when a defect in one of the membrane proteins of the nerve cell causes the subsequent accumulation of inert masses inside the cell and disruption of its vital functions.
Disruption of synthesis processes in a cell can occur at its various stages: RNA transcription in the nucleus, translation of polypeptides in ribosomes, post-translational modification, hypermethylation and glycosylation of the beige molecule, transport and distribution of proteins in the cell and their removal to the outside. In this case, one can observe an increase or decrease in the number of ribosomes, the breakdown of polyribosomes, expansion of the cisterns of the granular endoplasmic reticulum, loss of ribosomes, and the formation of vesicles and vacuoles. Thus, when poisoned by a pale grebe, the RNA polymerase enzyme is damaged, which disrupts transcription. Diphtheria toxin, by inactivating the elongation factor, disrupts translation processes, causing myocardial damage. The cause of disruption of the synthesis of some specific protein molecules can be infectious agents. For example, herpes viruses inhibit the synthesis and expression of MHC antigen molecules, which allows them to partially avoid immune control; plague bacilli - the synthesis of mediators of acute inflammation. The appearance of unusual proteins can stop their further breakdown and lead to the accumulation of inert or even toxic material. This can, to a certain extent, be facilitated by disruption of decay processes.
Disruption of decay processes
Simultaneously with the synthesis of protein in the cell, its breakdown continuously occurs. Under normal conditions, this has important regulatory and formative significance, for example, during the activation of inactive forms of enzymes, protein hormones, and mitotic cycle proteins. Normal cell growth and development require a finely controlled balance between the synthesis and degradation of proteins and organelles. However, in the process of protein synthesis, due to errors in the operation of the synthesizing apparatus, abnormal structuring of the protein molecule, and its damage by chemical and bacterial agents, a fairly large number of defective molecules are constantly formed. According to some estimates, their share is about a third of all synthesized proteins.
Mammalian cells have several main ways of protein destruction: through lysosomal proteases (pentide hydrolases), calcium-dependent proteinases (endopeptidases) and the proteasome system. In addition, there are also specialized proteinases, such as caspases. The main organelle in which degradation of substances occurs in eukaryotic cells is the lysosome, which contains numerous hydrolytic enzymes. Due to the processes of endocytosis and various types of autophagy in lysosomes and phagolysosomes, both defective protein molecules and entire organelles are destroyed: damaged mitochondria, sections of the plasma membrane, some extracellular proteins, and the contents of secretory granules.
An important mechanism for protein degradation is the proteasome, a multicatalytic proteinase structure of complex structure localized in the cytosol, nucleus, endoplasmic reticulum and on the cell membrane. This enzyme system is responsible for breaking down damaged proteins as well as healthy proteins that must be removed for normal cell function. In this case, the proteins to be destroyed are preliminarily combined with a specific polypeptide, ubiquitin. However, non-ubiquitinated proteins can also be partially destroyed in proteasomes. The breakdown of protein molecules in proteasomes into short polypeptides (processing) with their subsequent presentation together with type I MHC molecules is an important link in the immune control of antigenic homeostasis in the body. When proteasome function is weakened, damaged and unnecessary proteins accumulate, which accompanies cell aging. Violation of the degradation of cyclin-dependent proteins leads to disruption of cell division, degradation of secretory proteins - to the development of cystofibrosis. Conversely, an increase in proteasome function accompanies the depletion of the body (AIDS, cancer).
With genetically determined disorders of protein degradation, the organism is not viable and dies in the early stages of embryogenesis. If the breakdown of fats or carbohydrates is disrupted, storage diseases (thesaurismosis) occur. In this case, an excessive amount of certain substances or products of their incomplete breakdown - lipids, polysaccharides - accumulates inside the cell, which significantly damages the function of the cell. Most often this is observed in liver epithelial cells (hepatocytes), neurons, fibroblasts and macrophagocytes.
Acquired disorders of the processes of breakdown of substances can arise as a result of pathological processes (for example, protein, fat, carbohydrate and pigmentary degeneration) and are accompanied by the formation of unusual substances. Disturbances in the lysosomal proteolysis system lead to decreased adaptation during fasting or increased stress, and to the occurrence of certain endocrine dysfunctions - decreased levels of insulin, thyroglobulin, cytokines and their receptors. Impaired protein degradation slows down the rate of wound healing, causes the development of atherosclerosis, and affects the immune response. With hypoxia, changes in intracellular pH, radiation injury, characterized by increased peroxidation of membrane lipids, as well as under the influence of lysosomotropic substances - bacterial endotoxins, metabolites of toxic fungi (sporofusarin), silicon oxide crystals - the stability of the lysosome membrane changes, activated lysosomal enzymes are released into the cytoplasm , which causes destruction of cell structures and its death.
Chapter 1
BASICS OF CELL PHYSIOLOGY
I. Dudel
Plasma membrane . Animal cells are bounded by a plasma membrane (Figure 1.1). We will dwell on its structure, which is very similar to the structure of many intracellular membranes, in a little more detail. The main matrix of the membrane consists of lipids, mainly phosphatidylcholine. These lipids consist of a hydrophilic head group to which long hydrophobic hydrocarbon chains are attached. In water, such lipids spontaneously form a bilayer film 4–5 nm thick, in which the hydrophilic groups face the aqueous medium, and the hydrophobic hydrocarbon chains are arranged in two rows, forming an anhydrous lipid phase. Cell membranes are lipid bilayers of this type and contain glycolipids, cholesterol and phospholipids (Fig. 1.2). The hydrophilic part of glycolipids is formed by oligosaccharides. Glycolipids are always located on the outer surface of the plasma membrane, with the oligosaccharide part of the molecule oriented like a hair immersed in the environment. Scattered among the phospholipids in almost equal quantities, cholesterol molecules stabilize the membrane. The distribution of various lipids in the inner and outer layers of the membrane is not the same, and even within one layer there are areas in which certain types of lipids are concentrated. This uneven distribution
Rice. 1.1. Schematic drawing of a cell showing the most important organelles
probably has some, as yet unclear, functional significance.
The main functional elements embedded in the relatively inert lipid matrix of the membrane are squirrels(Fig. 1.2). Protein by mass accounts for 25 to 75% in various membranes, but since protein molecules are much larger than lipid molecules, 50% by mass is equivalent to the ratio of 1 protein molecule to 50 lipid molecules. Some proteins penetrate the membrane from its outer to inner surface, while others are fixed in one layer. Protein molecules are usually oriented so that their hydrophobic groups are embedded in the lipid membrane and the polar hydrophilic groups on the surface of the membrane are embedded in the aqueous phase. Many proteins on the outer surface of the membrane are glycoproteins; their hydrophilic saccharide groups face the extracellular environment.
Membrane systems of intracellular organelles .
Approximately half of the cell volume is occupied by organelles isolated from the cytosol by membranes. The total surface area of the membranes of intracellular organelles is at least 10 times greater than the surface of the plasma membrane. The most widely used membrane system is endoplasmic reticulum, representing a network

Rice. 1.2.Schematic representation of the plasma membrane. Proteins are embedded in the phospholipid bilayer, with some of them penetrating the bilayer, while others are only anchored to the outer or inner layer
highly convoluted tubes or sac-like elongated structures; large areas of the endoplasmic reticulum are studded with ribosomes; such a reticulum is called granular or rough (Fig. 1.1). Golgi apparatus also consists of membrane-bound lamellae, from which vesicles or vesicles break off (Fig. 1.1). Lysosomes and peroxisomes- These are small specialized vesicles. In all these various organelles, the membrane and the space it encompasses contain specific sets of enzymes; Inside the organelles, special metabolic products accumulate, used to carry out various functions of the organelles.
CoreAnd mitochondria differ in that each of these organelles is surrounded by two membranes. The nucleus is responsible for the kinetic control of metabolism; the folded inner membrane of the mitochondria is the site of oxidative metabolism; here, due to the oxidation of pyruvate or fatty acids, the high-energy compound adenosine triphosphate (ATP, or ATP) is synthesized.
Cytoskeleton . The cytoplasm surrounding the organelles cannot in any way be considered amorphous; it is permeated by a cytoskeletal network. The cytoskeleton consists of microtubules, actin filaments and intermediate filaments (Fig. 1.1). Microtubules have an outer diameter of about 25 nm; they are formed, like a regular polymer, as a result of the assembly of tubulin protein molecules. Actin filaments– contractile fibers, located in the near-membrane layer and throughout the cell, mainly take part in processes associated with movement. Intermediate filaments consist of blocks of different chemical composition in different types of cells; they form a variety of connections between the two other cytoskeletal elements mentioned above. Organelles and the plasma membrane are also associated with the cytoskeleton, which not only maintains the shape of the cell and the position of the organelles in it, but also determines changes in the shape of the cell and its mobility.
Cytosol . About half of the cell volume is occupied by the cytosol. Since it is approximately 20% protein by weight, it is more of a gel than an aqueous solution. Small molecules, including organic and inorganic ions, dissolved in the aqueous phase. An exchange of ions occurs between the cell and the environment (extracellular space); these metabolic processes will be discussed in the next section. The concentration of ions in the extracellular space is maintained with considerable accuracy at a constant level; the intracellular concentration of each ion also has a specific level, different from that outside the cell (Table 1.1). The most abundant cation in the extracellular environment is Na+ in the cell its concentration is more than 10 times lower. On the contrary, the concentration of K + is highest inside the cell; outside the cell it is lower by more than an order of magnitude. The greatest gradient between extracellular and intracellular concentrations exists for Ca 2+, the concentration of free ions inside the cell is at least 10,000 times lower than outside it. Not all ions are dissolved in the cytosol; some of them are adsorbed on proteins or deposited in organelles. For example, in the case of Ca 2+, bound ions are much more numerous than free ones. Most of the cytosolic proteins are enzymes, with the participation of which many processes of intermediate metabolism are carried out: glycolysis and gluconeogenesis, synthesis or destruction of amino acids, protein synthesis on ribosomes (Fig. 1.1). The cytosol also contains lipid droplets and glycogen granules, which serve as reserves of important molecules.
Table 1.1.Intra- and extracellular ion concentrations in muscle cells of homeothermic animals. A – – “high molecular weight cellular anions”
|
Intracellular concentration |
Extracellular concentration |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Others cations |
5 mmol/l |
|
Resting potential –90 mV |
|||
1.2. Exchange of substances between the cell and the environment
We have briefly described the structure of the cell so that we can use this description to review the basics of cellular physiology. In no case can a cell be considered a static entity, since there is a constant exchange of substances between various intracellular compartments, as well as between compartments and the environment. Cell structures are in dynamic equilibrium, and the interactions of cells with each other and with the external environment are a necessary condition for maintaining the life of a functioning organism. In this chapter we will look at the fundamental mechanisms of such exchange. In subsequent chapters these mechanisms will be considered as applied to the nerve cell and its functions;
however, the same mechanisms underlie the functioning of all other organs.
Diffusion.The simplest process of moving matter is diffusion. In solutions (or gases), atoms and molecules move freely, and differences in concentrations are balanced due to diffusion. Let's consider two volumes filled with liquid or gas (Fig. 1.3), in which substances have concentrations c 1 and c 2 and separated by a layer with surface area A and thickness d. Flow of matter m during time t described Fick's first law of diffusion:
dm/ dt= DA/ d ( C 1 –C 2)=DA/ dD C(1)
where D is the diffusion coefficient, constant for a given substance, solvent and temperature. In a more general form for the concentration difference dc at a distance dx
dm/dt= –D A dc/dx,(2)
the flow through section A is proportional to the concentration gradient dc/dx . The minus sign appears in the equation because the change in concentration in the x direction is negative.
Diffusion is the most important process due to which most of the molecules in aqueous solutions move over short distances. This also applies to their movement in the cell insofar as diffusion is not impeded by membranes. Many substances can diffuse freely through lipid membranes, especially water and dissolved gases such as O 2 and CO 2 . Fat soluble

Rice. 1.3.Quantitative diffusion scheme. The two spaces are separated by a layer of thicknessdand area A. C; – high concentration of particles in the left part of the volume, C: – low concentration of particles in the right parts, pink surface– concentration gradient in the diffusion layer. Diffusion flow dm/dt – see equation (1)
substances also diffuse well through membranes; This also applies to polar molecules of a fairly small size, such as ethanol and urea, while sugars pass through the lipid layer with difficulty. At the same time, lipid layers are practically impermeable to charged molecules, including even inorganic ions. For non-electrolytes, the diffusion equation (1) is usually transformed by combining the characteristics of the membrane and the diffusing substance into one parameter-permeability (P):
dm/dt=P AD c.(3)
In Fig. 1.4 are compared permeability (P) of the lipid membrane for various molecules.
Diffusion through membrane pores . The plasma membrane (and other cell membranes) is permeable not only to substances diffusing through the lipid layer, but also to many ions, sugars, amino acids and nucleotides. These substances cross the membrane through pores formed transport proteins, immersed in the membrane. Inside such proteins there is a water-filled channel with a diameter of less than 1 nm, through which small molecules can diffuse. They move along a concentration gradient, and if they carry a charge, then their movement through the channels is also regulated by the membrane potential. Membrane channels are relatively selective

Rice. 1.4.Permeability of artificial lipid bilayers to various substances
in relation to the type of molecules that can pass through them, there are, for example, potassium, sodium and calcium channels, each of which is impermeable to almost any ion except a specific one. Such selectivity is determined by the charge or structure of binding sites in the channel walls, which facilitates the transport of a specific molecule and prevents the penetration of other substances through the channel (Fig. 1.5, A) .
Behind the behavior membrane ion channels easy to observe, since the current arising from the movement of ions can be measured, even for a single channel. It is shown that the channels spontaneously and with high frequency change their state from open to closed. The potassium channel is characterized by current pulses with an amplitude of about 2 pA (2 10 –12 A) and a duration of several milliseconds (see Fig. 2.12, p. 37) [3]. During this period, tens of thousands of ions pass through it. The transition of proteins from one conformation to another is studied using X-ray diffraction, Mössbauer spectroscopy and nuclear magnetic resonance (NMR). Proteins are thus very dynamic, mobile structures, and the channel passing through the protein is not just a rigid, water-filled tube (Fig. 1.5, A), but a labyrinth of rapidly moving molecular groups and charges. This dynamic characteristic of the channel is reflected in energy profile of the channel, shown in Fig. 1.5, B. Here, the abscissa axis represents the length of the channel from the external solution with ion concentration C 0 and potential 0 to the internal solution with concentration C 1 and potential E. The ordinate axis

Rice. 1.5.A. Schematic of a protein forming a potassium channel embedded in the lipid bilayer of the plasma membrane. Four negative charges are fixed on the “wall” of the channel. B. Schematic energy profile of the channel shown in Fig. A. The ordinate axis shows the values of kinetic energy required for passage of the channel; along the abscissa axis – the distance between the inner and outer surfaces of the membrane. The energy minima correspond to the binding sites of positively charged ions with fixed negative charges in the channel wall. The energy maxima correspond to diffusion obstacles in the channel. The conformation of the channel protein is assumed to spontaneously oscillate; energy profile options are shown with solid and dashed lines; these oscillations greatly facilitate the binding of ions when overcoming the energy barrier (modified)
the ion energy levels at the channel binding sites are presented; the peak in the graph represents the permeability barrier that the ion energy must exceed to penetrate the channel, and the “dip” in the graph represents a relatively stable state (binding). Despite the obstacle of the energy peak, the ion can penetrate the channel if the energy profile changes spontaneously; the ion may thus suddenly find itself “on the other side” of the energy peak and may continue to move into the cell. Depending on the charge, size and degree of hydration of the ion and its ability to bind to channel wall structures, the energy profile of the channel varies for different ions, which may explain the selectivity of individual channel types.
Diffusion equilibrium of ions . The diffusion of various ions through membrane channels should lead to the elimination of differences in concentrations between the extra- and intracellular environments. As, however, can be seen from table. 1.1, such differences remain, therefore, there must be some equilibrium between diffusion and other transport processes across the membrane. The next two sections deal with the ways in which this equilibrium is achieved. In the case of ions, the diffusion equilibrium is affected by their charge. Diffusion of uncharged molecules is ensured by the difference in concentrations dc , and when the concentrations are equalized, the transport itself stops. Charged particles are additionally affected by the electric field. For example, when a potassium ion leaves a cell along its concentration gradient, it carries a single positive charge. Thus, the intracellular environment becomes more negatively charged, resulting in a potential difference across the membrane. The intracellular negative charge prevents new potassium ions from leaving the cell, and those ions that do leave the cell will further increase the charge on the membrane. The flow of potassium ions stops when the action of the electric field compensates for the diffusion pressure due to the difference in concentration. Ions continue to pass through the membrane, but in equal amounts in both directions. Therefore, for a given difference in ion concentrations on the membrane, there is equilibrium potential E ion at which the flow of ions through the membrane stops. The equilibrium potential can be easily determined using Nernst equations:
Eion= RT/ zF* lnC out/ C in(4)
where R – gas constant, T – absolute temperature, z – ion valence (negative for anions) C out – extracellular ion concentration, C in – intracellular ion concentration, F Faraday number. If we substitute constants into the equation, then at body temperature (T = 310 K) the equilibrium potential for potassium ions E K is equal to:
Ek= –61 mB log /(5)
If [ K + out ]/[ K + in ] = 39, as follows from table. 1.1 then
Ek= –61 m B log 39= –97 mV.
Indeed, it was found that all cells have membrane potential; in mammalian muscle cells its level is about -90 mV. Depending on conditions and relative ion concentrations, cells can have a membrane potential ranging from -40 to -120 mV. For the cell in the example above (Table 1.1) resting potential, equal to approximately -90 mV, indicates that the fluxes of potassium ions through membrane channels are approximately in equilibrium. This is not surprising, since in a resting membrane the open state of potassium channels is most likely, i.e. the membrane is most permeable to potassium ions. The membrane potential, however, is determined by the flows of other ions.
The ease with which uncharged particles can diffuse through the membrane is quantified in equation (3). Permeability to charged particles is described by a slightly more complex equation:
P= m RT/ dF(6)
Where m– ion mobility in the membrane, d – membrane thickness, a R, T and F – known thermodynamic constants. The permeability values for various ions determined in this way can be used to calculate the membrane potential Em , when potassium, sodium and chlorine ions pass through the membrane simultaneously (with permeability P K, P Na and P Cl respectively). It is assumed that the potential drops uniformly in the membrane, so that the field strength is constant. In this case it applies Goldman equation, or constant field equation :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
For most cell membranes P K approximately 30 times higher than P Na (see also section 1.3). Relative value P Cl varies greatly; for many membranes P Cl small compared to R K , however for others (for example, in skeletal muscles) P Cl , significantly higher than P K.
Active transport, sodium pump . The previous section describes the passive diffusion of ions and the resulting membrane potential at given intra- and extracellular ion concentrations. However, as a result of this process, the concentration of ions inside the cell is not automatically stabilized, since the membrane
potential is slightly more electronegative than EK, and much compared to E Na (about +60 mV). Due to diffusion, intracellular ion concentrations, at least potassium and sodium, must be equalized with extracellular ones. The stability of the ion gradient is achieved through active transport: membrane proteins transport ions across the membrane against electrical and/or concentration gradients, consuming metabolic energy for this. The most important process of active transport is the work Na/K – a pump that exists in almost all cells;
the pump pumps sodium ions out of the cell while simultaneously pumping potassium ions into the cell. This ensures a low intracellular concentration of sodium ions and a high concentration of potassium ions (Table 1.1). The concentration gradient of sodium ions on the membrane has specific functions related to the transmission of information in the form of electrical impulses (see Section 2.2), as well as the maintenance of other active transport mechanisms and regulation of cell volume (see below). Therefore, it is not surprising that more than 1/3 of the energy consumed by a cell is spent on the Na/K pump, and in some of the most active cells up to 70% of energy is spent on its operation.
The Na/K transport protein is an ATPase. On the inner surface of the membrane, it breaks down ATP into ADP and phosphate (Fig. 1.6). The energy of one ATP molecule is used to transport three sodium ions from the cell and simultaneously two potassium ions into the cell, i.e., in total, one positive charge is removed from the cell in one cycle. Thus, the Na/K pump is electrogenic(creates an electrical current across the membrane), which leads to an increase in electronegativity of the membrane potential by approximately 10 mV. The transport protein performs this operation at high speed: from 150 to 600 sodium ions per second. The amino acid sequence of the transport protein is known, but the mechanism of this complex metabolic transport is not yet clear. This process is described using energy profiles of the transfer of sodium or potassium ions by proteins (Fig. 1.5,5). By the nature of the changes in these profiles, associated with constant changes in the conformation of the transport protein (a process that requires energy), one can judge the stoichiometry of the exchange: two potassium ions are exchanged for three sodium ions.
Na/K pump, same as isolated Na+ /K + -dependent membrane ATPase, specifically inhibited by the cardiac glycoside ouabain (strophanthin). Since the operation of the Na/K pump is a multi-stage chemical reaction, it, like all chemical reactions, is largely dependent on temperature, which

Rice. 1.6.Diagram of a Na/K pump-ATPase (immersed in the lipid bilayer of the plasma membrane), which in one cycle removes three Na + ions from the cell against potential and concentration gradients and brings two K ions into the cell + . During this process, one ATP molecule is broken down into ADP and phosphate. The diagram shows the ATPase as a dimer consisting of a large (functional) and small subunits; in the membrane it exists as a tetramer formed by two large and two small subunits
shown in Fig. 1.7. Here, the flux of sodium ions from muscle cells is shown relative to time; this is practically equivalent to the flow of sodium ions mediated by the operation of the Na/K pump, because the passive flow of sodium ions against concentration and potential gradients is extremely small. If the drug is cooled by approximately 18°C, the flow of sodium ions from the cell will quickly decrease by 15 times, and immediately after heating it will be restored to its original level. This decrease in the flow of sodium ions from the cell is several times greater than that which would correspond to the temperature dependence of the diffusion process or a simple chemical reaction. A similar effect is observed when metabolic energy is depleted as a result of dinitrophenol (DNP) poisoning (Fig. 1.7,5). Consequently, the flow of sodium ions from the cell is ensured by an energy-dependent reaction - an active pump. Another characteristic of the pump, along with significant temperature and energy dependence, is the presence of a saturation level (like all other chemical reactions); this means that the speed of the pump cannot increase indefinitely as the concentration of transported ions increases (Fig. 1.8). In contrast, the flux of a passively diffusing substance increases in proportion to the concentration difference in accordance with the law of diffusion (equations 1 and 2).

Rice. 1.7. A, B. Active Na transport + . Y axis: flux of radioactive 24 Na + from the cell (imp./min). Abscissa axis: time since the start of the experiment. A. The cell is cooled from 18.3 °C to 0.5 °C; flow Na+ from the cell during this period is inhibited. B. Suppression of Na + flux from the cell with dinitrophenol (DNP) at a concentration of 0.2 mmol/l (modified)
In addition to the Na/K pump, the plasma membrane contains at least one more pump - calcium; this pump pumps calcium ions (Ca 2+) out of the cell and is involved in maintaining their intracellular concentration at an extremely low level (Table 1.1). The calcium pump is present in very high density in the sarcoplasmic reticulum of muscle cells, which accumulate calcium ions as a result of the breakdown of ATP molecules (see Chapter 4).
Effect of the Na/K pump on membrane potential and cell volume . In Fig. Figure 1.9 shows the various components of the membrane current and shows the intracellular concentrations of ions that

Rice. 1.8.The relationship between the rate of transport of molecules and their concentration (at the entrance to the channel or at the binding site of the pump) during diffusion through the channel or during pumping transport. The latter becomes saturated at high concentrations (maximum speed, Vmax ) value on the x-axis corresponding to half the maximum pump speed ( Vmax /2), is the equilibrium concentration TO m

Rice. 1.9.Diagram showing Na+ concentrations , K+ and Cl – inside and outside the cell and the pathways for the penetration of these ions through the cell membrane (through specific ion channels or using a Na/K pump. At given concentration gradients, equilibrium potentials E Na, E K and E С l – equal to those indicated, membrane potential Em = – 90 mV
ensure their existence. An outward current of potassium ions is observed through potassium channels, since the membrane potential is slightly more electropositive than the equilibrium potential for potassium ions. The overall conductance of sodium channels is much lower than that of potassium channels, i.e. sodium channels are open much less frequently than potassium channels at resting potential; however, approximately the same number of sodium ions enter the cell as potassium ions exit the cell, because large concentration and potential gradients are required for sodium ions to diffuse into the cell. The Na/K pump provides ideal compensation for passive diffusion currents, as it transports sodium ions out of the cell and potassium ions into it. Thus, the pump is electrogenic due to the difference in the number of charges transferred into and out of the cell, which at normal speed of its operation creates a membrane potential of approximately 10 mV is more electronegative than if it were produced by passive ion fluxes alone (see Equation 7). As a result, the membrane potential approaches the potassium equilibrium potential, which reduces the leakage of potassium ions. Na activity The /K pump is regulated by the intracellular concentration of sodium ions. The speed of the pump slows down as the concentration of sodium ions to be removed from the cell decreases (Fig. 1.8), so that the pump operation and the flow of sodium ions into the cell balance each other, maintaining the intracellular concentration of sodium ions at a level of approximately 10 mmol/L.
To maintain equilibrium between pumping and passive membrane currents, many more Na/K pump molecules are needed than channel proteins for potassium and sodium ions. When the channel is open, tens of thousands of ions pass through it in a few milliseconds (see above), and since the channel usually opens several times per second, in total more than 10 5 ions pass through it during this time. A single pump protein moves several hundred sodium ions per second, so the plasma membrane must contain about 1000 times more pump molecules than channel molecules. Measurements of channel currents at rest showed the presence on average of one potassium and one sodium open channel per 1 μm 2 membrane; It follows from this that about 1000 molecules of the Na/K pump should be present in the same space, i.e. the distance between them is on average 34 nm; The diameter of the pump protein, as a channel protein, is 8–10 nm. Thus, the membrane is quite densely saturated with pumping molecules
The fact that the flow of sodium ions into the cell, and potassium ions out of the cell, is compensated by the operation of the pump, there is another consequence, which consists in maintaining a stable osmotic pressure and constant volume. Inside the cell there is a high concentration of large anions, mainly proteins (A - in Table 1.1), which are not able to penetrate the membrane (or penetrate through it very slowly) and therefore are a fixed component inside the cell. To balance the charge of these anions, an equal number of cations is needed. Due to the action of the Na/K pump, these cations are mainly potassium ions. A significant increase in the intracellular ion concentration could only occur with an increase in the concentration of anions due to the flow of C1 - along the concentration gradient into the cell (Table 1.1), but the membrane potential counteracts this. Incoming current Cl – observed only until the equilibrium potential for chlorine ions is reached; this is observed when the chlorine ion gradient is almost opposite to the potassium ion gradient, since chlorine ions are negatively charged (Equation 4). Thus, a low intracellular concentration of chlorine ions is established, corresponding to a low extracellular concentration of potassium ions. The result is a limitation of the total number of ions in the cell. If the membrane potential drops when the Na/K pump is blocked, for example during anoxia, then the equilibrium potential for chlorine ions decreases, and the intracellular concentration of chloride ions increases accordingly. Restoring the balance of charges, potassium ions also enter the cell; the total concentration of ions in the cell increases, which increases osmotic pressure; this forces water into the cell. The cell swells. This swelling is observed in vivo in conditions of lack of energy.
Concentration gradient Na + as a driving force for membrane transport . The importance of the Na/K pump for the cell is not limited to the stabilization of normal K + and Na + gradients on the membrane. Energy stored in membrane gradient Na+ , is often used to facilitate membrane transport of other substances. For example, in Fig. 1.10 shows “simport” Na+ and sugar molecules into the cell. The membrane transport protein transports the sugar molecule into the cell even against a concentration gradient, while at the same time Na + moves along the concentration and potential gradient, providing energy for transport of sugars. This transport of sugars is entirely dependent on the existence of a high gradient Na+ ; if intracellular concentration Na+ increases significantly, the transport of sugars stops. For various c akharov, there are different simport systems. Amino acid transport into a cell similar to transport c akharov shown in Fig. 1.10; it is also provided by the gradient Na+ , There are at least five different symport systems, each specialized for one group of related amino acids.
In addition to simport systems, there are also "anti-porters". One of them, for example, transfers one calcium ion out of the cell in one cycle in exchange for three incoming sodium ions (Fig. 1.10). Energy for Ca 2+ transport is formed due to the entry of three sodium ions along a concentration and potential gradient. This energy is sufficient (at resting potential) to maintain a high calcium ion gradient (from less than 10–7 mol/L inside the cell to approximately 2 mmol/L outside the cell).
Endo- and exocytosis . For certain substances that enter the cell or must be excreted

Rice. 1.10.Proteins embedded in the lipid bilayer of the membrane mediate the symport of glucose and Na + into the cell, as well as Ca2+/Na+ –antiport, in which the driving force is the Na + gradient on the cell membrane
from it, there are no transport channels; These substances include, for example, proteins and cholesterol. They can pass through the plasma membrane into vesicles, or bubbles, through endo- and exocytosis. In Fig. Figure 1.11 shows the main mechanisms of these processes. In exocytosis, certain organelles (see below) form vesicles filled with a substance that needs to be removed from the cell, such as hormones or extracellular enzymes. When such vesicles reach the plasma membrane, their lipid membrane fuses with it, thus allowing the contents to escape into the external environment. In the opposite process, endocytosis, the plasma membrane invaginates, forming a pit, which then deepens and closes, forming an intracellular vesicle filled with extracellular fluid and some macromolecules. To ensure this membrane fusion and closure of the vesicle, the contractile elements of the cytoskeleton act in conjunction with the membranes themselves (see below). Endocytosis does not always simply involve the capture of the extracellular environment into the cell. The cell membrane often contains specific receptors for macromolecules, such as insulin or antigens, often organized into specialized groups. After these macromolecules bind to their receptors, endocytosis occurs in the membrane area surrounding the receptor, and the macromolecule is selectively transported into the cell (Fig. 1.12, B).
Endo- and exocytosis occur continuously in cells. The amount of membrane material turnover is significant; within 1 hour, the macrophage absorbs double the surface area of its cytoplasmic membrane in the form of vesicles. In most cells, the turnover of membrane material does not occur as intensively, but it should still be significant.

Rice. 1.11.Exocytosis and endocytosis. Up: the intracellular vesicle fuses with the lipid bilayer of the plasma membrane and opens into the extracellular space. This process is called exocytosis. At the bottom: the plasma membrane invaginates in a small area and releases a vesicle filled with extracellular material. This process is called endocytosis
1.3. Transport of substances within the cell
Endo- and exocytosis are not only processes of transport of substances across the cell membrane, but also processes of exchange of membranes - the structural components of the cell itself. The subject of consideration in this section are other similar transport processes in the cell and its organelles.

Rice. 1.12. A–B. Scheme of processes including exo- and endocytosis. A. The protein synthesized in the granular endoplasmic reticulum is transported through the Golgi apparatus to the plasma membrane, where it is secreted by exocytosis. B. Cholesterol bound to LDL (low-density lipoprotein) particles attaches to the plasma membrane, induces the formation of an endocytic vesicle in this region of the membrane and is transported to lysosomes, where it is released. IN. Extracellular material captured during endocytosis (in the figure on right), transported through the cell in vesicles, or vesicles, and released through exocytosis (in the figure left)
Diffusion . Naturally, in the cytosol the difference in concentrations is eliminated due to diffusion; the same is true for the fluids contained in the organelles. Due to the high concentration of dissolved protein, diffusion here is much slower than in water. Lipid membranes—around cells and within organelles—are two-dimensional fluids in which diffusion occurs. Lipids in a membrane bilayer diffuse within their own layer, rarely moving from one to another. The proteins immersed in them are also quite mobile; they rotate around an axis perpendicular to the membrane or diffuse laterally with very different diffusion constants, 2–10,000 times slower than phospholipids. So, if some proteins move freely in the lipid layer and at the same speed as the lipid molecules themselves, then others are anchored, i.e. quite firmly connected to the cytoskeleton. There are “permanent” aggregates of specific proteins in the membrane, for example, pre- and postsynaptic structures of nerve cells. Freely moving proteins can be demonstrated by binding them to fluorescent dyes, the luminescence of which is induced by briefly illuminating a small area of the membrane. Such experiments show that in less than 1 min, proteins bound to the dye are uniformly distributed over the membrane over distances of up to 10 μm.
Active transport in organelle membranes .
Active transport processes, which play a vital role in the functioning of the plasma membrane, also occur inside the cell, in the membranes of organelles. The specific contents of various organelles are created partly due to internal synthesis, and partly due to active transport from the cytosol. One example of the latter is the above-mentioned Ca 2+ pump in the sarcoplasmic reticulum of muscle cells. It is especially interesting that in the case of ATP synthesis in mitochondria, the opposite principle applies to what occurs in the ATPase pumps of the plasma membrane (Fig. 1.6). During ATP synthesis, oxidative metabolism leads to the formation of a steep gradient H+ on internal membranes. This gradient is the driving force for the reverse process of the pumping cycle of active transport of molecules: H + ions move across the membrane along the gradient, and the energy released as a result provides the synthesis of ATP from ADP and phosphate. The resulting ATP, in turn, provides energy to the cell, including for active transport.
Transport in vesicles . The cell has a large number of organelles and associated vesicles (Fig. 1.1). These organelles, and especially the vesicles, are in constant motion, transporting their contents to other organelles or to the plasma membrane. Vesicles can also migrate from the cell membrane to organelles, as in endocytosis.
Process protein secretion shown in Fig. 1.12, A. The protein is synthesized near the cell nucleus on ribosomes associated with the endoplasmic reticulum (the so-called granular, or rough, endoplasmic reticulum); Once in the endoplasmic reticulum, the protein is packaged into transport vesicles, which separate from the organelle and migrate to the Golgi apparatus. Here they merge with the cisternae of the Golgi apparatus, where the protein is modified (i.e., converted into a glycoprotein). At the ends of the cisternae, the vesicles separate again. Secretory vesicles carrying the modified protein move to the plasma membrane and release their contents by exocytosis.
Another example of a transport pathway in a cell is shown in Fig. 1.12, B; This is the absorption of cholesterol by the cell. Cholesterol transported in the blood is mainly bound to proteins, such as particles "low density lipoprotein"(LDL). Such particles attach to specific membrane areas containing LDL receptors, where endocytosis occurs and LDL is transported into the cell in “bordered” vesicles. These vesicles fuse to form endosomes and lose their “edging” during this process. Endosomes, in turn, merge with primary lysosomes, containing predominantly hydrolytic enzymes, and form secondary, larger lysosomes. In them, cholesterol is released from LDL particles and diffuses into the cytosol, where it becomes available, for example, for the synthesis of lipid membranes. Vesicles that do not contain LDL are also separated from endosomes, which move in a special way to the plasma membrane and merge with it, returning membrane material and, probably, receptors for LDL. From the moment the LDL particle binds to the membrane until cholesterol is released from the secondary lysosome, 10-15 minutes pass. Impairments in the binding and absorption of LDL, i.e., in the supply of cholesterol to the cell, play a decisive role in the development of a serious and widespread disease, atherosclerosis (“hardening” of the arteries).
There are many other transport routes similar to those shown in Fig. 1.11 and 1.12, A, with the help of which specific vesicles move in the cell. It is not known exactly how they move, but cytoskeletal elements are probably involved in this process. Vesicles can slide along microtubules, in which case the energy for movement is apparently provided by the protein associated with the vesicles, ATPase (see below). It remains completely unclear how many different vesicles, moving one after another in all directions, reach their destination. They obviously need to be “marked” in a way that is recognized by the transport system and converted into targeted movement.
Transport by formation and destruction of organelles . So far, we have considered endo- and exocytosis as processes of transporting the contents of vesicles. There is another aspect of these processes, which lies in the fact that the targeted removal of the plasma membrane on one part of the cell surface by endocytosis and, on the contrary, its addition on another by exocytosis moves large areas of the membrane (Fig. 1.12.E), giving the cell the opportunity, for example , form an outgrowth or move.
Similar rearrangements are also typical for the cytoskeleton, especially for microfilaments and microtubules (Fig. 1.1). Microfilaments consist primarily of F-actin protein, which is capable of assembly into fibrous bundles as a result of polymerization of the monomer from the cytosol. The bundles are polarized, that is, they often grow only at one end, accumulating new actin molecules, while the other end is inert or disassembly occurs here. Due to this polarized growth, microfilaments move efficiently and the structure of their network can change. The transition of actin from a depolymerized state (sol) to an organized state (gel) can occur very quickly under the influence of other proteins or changes in ion concentration (see below). There are also proteins that cause actin filaments to break down to form short fragments. The thin projections of many cells - filopodia - contain a central bundle of actin (Fig. 1.1), and the various movements of the filopodia are probably due to actin transitions: polymerization - depolymerization.
Microtubulesalso often undergo similar movements. The mechanism of these movements is similar - polymerization of tubulin from the cytosol in such a way that one of the ends of the microtubule grows, while the other either does not change, or disassembly occurs there. Thus, a microtubule, by appropriate addition or removal of material, can move throughout the cytosol.
Active movements of the cytoskeleton . Changes in cytoskeletal structures can occur as a result of both active movements and rearrangements described above. In many cases, the movement of microtubules and actin filaments is driven by contractile proteins that bind the filaments or tubules and can move them relative to each other. Squirrels myosin and dynein present in the cytosol of all cells in relatively high concentrations; they are the elements that convert energy into movement in specialized cells (muscle) and organelles (cilia). In muscle cells, myosin forms thick filaments oriented parallel to actin filaments. The myosin molecule with its “head” attaches to the actin filament and, using the energy of ATP, displaces myosin along the actin molecule. Myosin then detaches from actin. The combination of many such connection-disconnection cycles leads to macroscopic contraction of muscle fibers(chapter 4). Dynein plays a similar role in the movement of microtubules during cilia (Fig. 1.1). In the cytoplasm of unspecialized cells, myosin and dynein do not form regular fibers, but in most cases small groups of molecules. Even in such small aggregates, they are capable of moving actin filaments or microtubules. Rice. Figure 1.13 illustrates this process when oppositely polarized myosin molecules are also attached to two actin filaments, polarized in different directions. The myosin head groups bend toward the tail of the molecule, consuming ATP in the process, and the two actin filaments move in the opposite direction, after which the myosin detaches from them. Movements of this kind, during which ATP energy is converted into mechanical work, can change the shape of the cytoskeleton and, consequently, the cell, and also provide transport of organelles associated with the cytoskeleton.
The processes of intracellular transport can be most clearly demonstrated on the axon of a nerve cell. Axon transport is discussed here in detail to illustrate events that are likely to occur in similar ways in most cells. An axon that is only a few microns in diameter can reach a length of one meter or more, and the movement of proteins by diffusion from the nucleus to the distal end of the axon would take years. It has long been known that when any part of the axon undergoes constriction, the part of the axon located more proximally expands. It looks as if centrifugal flow is blocked in the axon. Such flow–fast axon transport can be demonstrated by the movement of radioactive markers, as in the experiment shown in Fig. 1.14. Radiolabeled leucine was injected into the dorsal root ganglion, and then radioactivity was measured in the sciatic nerve at a distance of 166 mm from the neuronal cell bodies from the 2nd to the 10th hour. Over 10 hours, the peak of radioactivity at the injection site changed slightly. But the wave of radioactivity spread along the axon at a constant speed of about 34 mm in 2 hours, or 410 mm/day. It has been shown that in all neurons of homeothermic animals, fast axonal transport occurs at the same speed, and no noticeable differences are observed between thin, unmyelinated fibers and the thickest axons, as well as between motor and sensory fibers. The type of radioactive marker also does not affect the rate of fast axonal transport; markers can serve as a variety of radioactive

Rice. 1.13.The non-muscle myosin complex, with a certain orientation, can bind to actin filaments of different polarity and, using the energy of ATP, displace them relative to each other
molecules, such as various amino acids, that are included in the proteins of the cell body of the neuron. If we analyze the peripheral part of the nerve to determine the nature of the carriers of radioactivity transported here, then such carriers are found mainly in the protein fraction, but also in the composition of mediators and free amino acids. Knowing that the properties of these substances are different and the sizes of their molecules are especially different, we can explain the constant speed of transport only by a transport mechanism common to all of them.
Described above fast axon transport is anterograde, i.e., directed away from the cell body. It has been shown that some substances move from the periphery to the cell body using retrograde transport. For example, acetylcholinesterase is transported in this direction at a speed 2 times slower than the speed of fast axonal transport. A marker often used in neuroanatomy, horseradish peroxidase, also moves by retrograde transport. Retrograde transport probably plays an important role in the regulation of protein synthesis in the cell body. A few days after the axon is cut, chromatolysis is observed in the cell body, which indicates a disruption of protein synthesis. The time required for chromatolysis correlates with the duration of retrograde transport from the site of axon transection to the cell body. This result also suggests an explanation for this disorder - the transmission from the periphery of the “signal substance” that regulates protein synthesis is disrupted. It is obvious that the main "vehicles" used for fast axonal
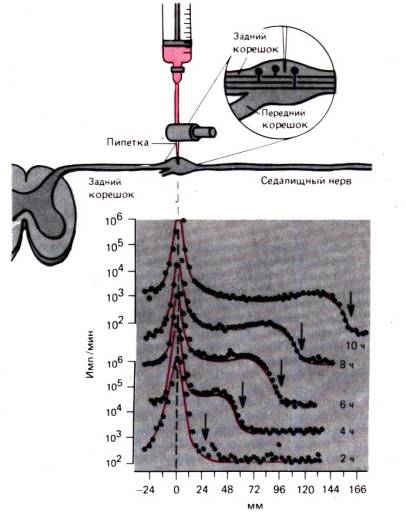
Rice. 1.14.Experiment demonstrating rapid axonal transport in sensory fibers of the cat sciatic nerve. Tritium-labeled leucine is injected into the dorsal root ganglion and radioactivity in the ganglion and sensory fibers is measured at 2, 4, 6, 8 and 10 hours after injection. (bottom of the picture). By x-axis The distance from the ganglion to the areas of the sciatic nerve where the measurement is made is plotted. On the ordinate axis only for the upper and lower curves, radioactivity (pulses/min) is plotted on a logarithmic scale. "Wave" of increased radioactivity (arrows) moves at a speed of 410 mm/day (along)
transport are vesicles (vesicles) and organelles, such as mitochondria, which contain substances that need to be transported. The movement of the largest vesicles or mitochondria can be observed using a microscope in vivo . Such particles make short, rapid movements in one direction, stop, often move slightly backward or to the side, stop again, and then jerk in the main direction. 410 mm/day corresponds to an average speed of anterograde movement of approximately 5 μm/s; the speed of each individual movement must, therefore, be much higher, and if we take into account the sizes of organelles, filaments and microtubules, then these movements are indeed very fast. Fast axonal transport requires a significant concentration of ATP. Poisons such as colchicine, which disrupts microtubules, also block fast axonal transport. It follows from this that in the transport process we are considering, vesicles and organelles move along microtubules and actin filaments; this movement is provided by small aggregates of dynein and myosin molecules, acting as shown in Fig. 1.13, using ATP energy.
Fast axon transport may also be involved in pathological processes. Some neurotropic viruses (for example, herpes or polio viruses) enter the axon at the periphery and move via retrograde transport to the neuron body, where they multiply and exert their toxic effects. Tetanus toxin, a protein produced by bacteria that enter the body when the skin is damaged, is captured by nerve endings and transported to the neuron body, where it causes characteristic muscle spasms. There are known cases of toxic effects on axon transport itself, for example, exposure to the industrial solvent acrylamide. In addition, it is believed that the pathogenesis of vitamin deficiency beriberi and alcoholic polyneuropathy involves a violation of fast axonal transport.
In addition to fast axonal transport in the cell, there is also quite intense slow axon transport. Tubulin moves along the axon at a speed of about 1 mm/day, and actin moves faster—up to 5 mm/day. Other proteins migrate with these cytoskeletal components; for example, the enzymes appear to be associated with actin or tubulin. The rates of movement of tubulin and actin are roughly consistent with the growth rates found for the mechanism described previously, where the molecules are incorporated into the active end of a microtubule or microfilament. Therefore, this mechanism may underlie slow axonal transport. The rate of slow axonal transport also approximately corresponds to the rate of axon growth, which apparently indicates restrictions imposed by the structure of the cytoskeleton on the second process.
Concluding this section, it should be emphasized that cells are by no means static structures, as they appear, for example, in electron microscopic photographs. Plasma membrane and especially organelles are in constant rapid movement and constant restructuring; that is why they are able to function. Further, these are not simple chambers in which chemical reactions take place, but highly organized conglomerates of membranes and fibers, in which reactions occur in an optimally organized sequence.
1.4. Regulation of cellular functions
The maintenance of the individual cell as a functional unit is primarily regulated by the nucleus; the study of such regulatory mechanisms is the subject of cell biology and biochemistry. At the same time, cells must modify their functions in accordance with environmental conditions and the needs of other cells of the body, i.e. they serve as objects of functional regulation. Below we will briefly consider how these regulatory influences act on the plasma membrane and how they reach intracellular organelles.
Regulatory effects on the cell membrane
Membrane potential . In many cases, the regulation of cellular functions is carried out by changing the membrane potential. Local changes in potential are possible when: 1) current from an adjacent area of the cell or generated by another cell flows through the membrane; 2) the extracellular concentration of ions changes (often [K + ] out ); 3) membrane ion channels open. Changes in membrane potential can affect the conformation of membrane proteins, causing, in particular, channels to open or close. As described above, the functioning of some membrane pumps depends on the membrane potential. Nerve cells are specialized to perceive changes in membrane potential as information that must be processed and transmitted (see Chapter 2).
Extracellular regulatory substances . The most important regulatory mechanism involving extracellular substances is their interaction with specific receptors on the plasma membrane or inside the cell. These substances include synaptic transmitters, which transmit information between nerve cells, local agents, and substances that circulate in the blood and reach all cells of the body, such as hormones and antigens. Synaptic transmitters are small molecules released from nerve endings at the synapse;
when they reach the plasma membrane of a neighboring, postsynaptic cell, they trigger electrical signals or other regulatory mechanisms. This issue is discussed in detail in Chap. 3.
Local chemical agents often secreted by specialized cells. They diffuse freely in the extracellular space, but their action is limited to a small group of cells due to the rapid destruction of these substances, either spontaneously or under the action of enzymes. One example of the release of such agents is the release histamine mast cells due to damage or immune response. Histamine causes relaxation of vascular smooth muscle cells, increases the permeability of the vascular endothelium and stimulates sensory nerve endings that mediate the sensation of itching. Other local chemical agents are released by many other cells. Typical local agents are prostaglandins, making up a group of approximately 20 fatty acid derivatives. They are released continuously from widely distributed cells, but act only locally, as they are quickly destroyed by membrane phospholipases. Various prostaglandins have a wide spectrum of action: they can trigger the contraction of smooth muscle cells, cause aggregation of blood platelets (platelets), or suppress the development of the corpus luteum in the ovaries.
Other local agents serve growth factors. The best known is nerve growth factor (NGF) for sympathetic neurons, which is necessary for the growth and survival of these neurons during development in vivo or in cell culture. Apparently, the target cells for this class of neurons secrete NGF and thereby ensure proper innervation. When forming organs, cells often need to “find their way” to target cells, which can be located at considerable distances. Accordingly, there must be many specialized growth factors similar to NGF.
Hormones and antigens carried by blood to all cells. Antigens trigger an immune response from cells bearing specific antibodies. However, antigens, as a rule, are foreign substances that are not produced in the reacting organism (for more details, see Chapter 18). Some hormones, such as insulin or thyroxine, affect a wide variety of cell types, while others, such as sex hormones, affect only a specific type of cell. Hormones are either peptides, whose action is triggered by binding to a receptor on the cell membrane, or steroids and thyroxine, which diffuse through the lipid membrane and bind to intracellular receptors. Steroid hormones bind to nuclear chromatin, resulting in the transcription of certain genes. The resulting proteins cause changes in cellular functions, which is the specific effect of hormones. Issues related to the release and action of hormones are discussed in detail in Chapter. 17.
Intracellular communication involving second messengers
The regulatory functions described above include effects on the cell membrane. Information received by the cell membrane must often cause organelles to react and is transferred to them by various substances known as second messengers (as opposed to first messengers, which come to the cell from external sources). The study of second mediators is evolving rapidly, and there is no guarantee that the current level of understanding of the problem will be sufficiently complete. Here we will touch on three well-studied mediators: Ca 2+ , cAMP and inositol triphosphate.
Calcium.The simplest intracellular mediator is the Ca 2+ ion. Its free concentration in a resting cell is very low and amounts to 10_–8–10–7 mol/l. It can enter the cell through specific membrane channels when they are in an open state, for example, when the membrane potential changes (see Chapter 2). The resulting increase in Ca 2+ concentration triggers important reactions in the cell, such as the contraction of myofibrils, which is the basis of muscle contraction (see Chapter 4), or the release of vesicles containing neurotransmitters from nerve endings (see Chapter 3) . Both reactions require a Ca 2+ concentration of approximately 10 –5 mol/l. Ca 2+, which has a regulatory effect, can also be released from intracellular stores, such as the endoplasmic reticulum. The release of Ca 2+ from the depot requires the participation of other intermediaries (see, for example, Fig. 1.16).
Cyclic adenosine monophosphate, cAMP. Recently, it has been proven that cyclic adenosine monophosphate (cAMP), a derivative of the body's main energy source, ATP, is an important second messenger. The complex chain of reactions shown in Fig. 1.15, starts with receptor R s on the outer surface of the plasma membrane, which can serve as a specific binding site for various mediators and hormones. After binding to a specific "stimulating" molecule R s changes its conformation; these changes affect the protein G s on the inner surface of the membrane in such a way that activation of the latter by intracellular guanosine triphosphate (GTP) becomes possible. Activated protein G s , in turn, stimulates the enzyme on the inner surface of the membrane, adenylate cyclase (AC), which catalyzes the formation of cAMP from ATP. Water-soluble cAMP is the mediator that transmits the effect

Rice. 1.15.A chain of reactions involving the intracellular messenger cAMP (cyclic adenosine monophosphate). Excitatory or inhibitory external signals activate membrane receptors R s or Ri . These receptors regulate the binding process G – proteins with intracellular GTP (guanosine triphosphate), thereby stimulating or inhibiting intracellular adenylate cyclase (AC). The amplifying enzyme AC converts adenosine triphosphate (ATP) into cAMP, which is then broken down into AMP by phosphodiesterase. Free cAMP diffuses into the cell and activates adenylate kinase (A-kinase), releasing its catalytic subunit C, which catalyzes the phosphorylation of intracellular proteins, i.e. forms the final effect of an extracellular stimulus. The diagram also shows pharmacological drugs and toxins that trigger (+) or inhibit (-) certain reactions (modified)
extracellular receptor stimulation R s to the internal structures of the cell.
In parallel with the stimulatory chain of reactions involving R s binding of inhibitory mediators and hormones to the corresponding receptor is possible R i which again through GTP-activated protein G , inhibits AC and thus the production of cAMP. Diffusing into the cell, cAMP reacts with adenylate kinase (A-kinase); this releases subunit C, which catalyzes the phosphorylation of protein P. This phosphorylation converts proteins into an active form, and they can now exert their specific regulatory effects (for example, cause glycogen degradation). This complex regulatory system is extremely efficient because the end result is the phosphorylation of many proteins, i.e., the regulatory signal passes through the circuit with a high gain. External mediators that bind to receptors R s and R i specific to each of them, extremely diverse. Adrenaline, connecting with R s or R i participates in the regulation of lipid and glycogen metabolism, as well as in enhancing the contraction of the heart muscle and in other reactions (see Chapter 19). Thyroid-stimulating hormone, activating R s , stimulates the release of the hormone thyroxine by the thyroid gland, and prostaglandin I inhibits the aggregation of blood platelets. Inhibitory effects, including adrenaline, mediated through R i are expressed in slowing down lipolysis. Thus, the cAMP system is a multifunctional intracellular regulatory system, which can be precisely controlled by extracellular stimulatory and inhibitory signaling substances.
Inositol phosphate "IF" h " The intracellular system of the second messenger, inositol phosphate, was discovered only recently (Fig. 1.16). In this case, there is no inhibitory pathway, but there is a similarity to the cAMP system, in which the effect of stimulation of the R receptor is transferred to the GTP-activated G protein on the inner surface of the membrane. At the next stage, the usual membrane lipid phosphatidylinositol (PI), having previously received two additional phosphate groups, is converted into PI-diphosphate (PIP 2), which is cleaved by activated phosphodiesterase (PDE) into inositol triphosphate(IFz) and lipid diacylglycerol(DAG). Inositol triphosphate is a water-soluble second messenger that diffuses into the cytosol. It acts primarily by releasing Ca 2+ from the endoplasmic reticulum. Ca 2+ in turn acts as a mediator, as described above; for example, it activates Ca 2+ -dependent phosphokinase, which phosphorylates enzymes. The lipid subunit of DAG (Fig. 1.16) also carries the signal, diffusing in the lipid phase of the plasma membrane to C-kinase located on its inner surface, which is activated with the participation of phosphatidylserine as a cofactor. Then C-kinase triggers phosphorylation of proteins, converting them into an active form.
The intracellular second messenger system IF 3 can also be controlled by a variety of external mediators and hormones, including acetylcholine, serotonin, vasopressin, and thyroid-stimulating hormone; like the cAMP system, it is characterized by a variety of intracellular effects. It is possible that this system is also activated by light in the visual receptor of the eye and plays a central role in phototransduction (see Chapter 11). For the first time in the individual development of the organism, the receptor of the IG system is activated by sperm, as a result of which IG takes part in the regulatory reactions accompanying the fertilization of the egg.
The cAMP and IFz-DAG systems are highly effective biological enhancers. They

Rice. 1.16.A chain of reactions involving the intracellular mediator IPG (inositol triphosphate). As in the cAMP system, the extracellular signal is mediated through a proteinG, which in this case activates phosphodiesterase (PDE). This enzyme breaks down phosphatidylinosine diphosphate (PIP 2 ) in the plasma membrane before IF h and diacylglycerol (DAG); IF h diffuses into the cytoplasm. Here it causes the release of Ca 2+ from the endoplasmic reticulum; increase in Ca concentration 2+ in the cytoplasm ([Ca 2+ ]i ) activates protein kinase, which phosphorylates and therefore activates enzymes. Another product, DAG, remains in the membrane and activates protein kinase C (phosphatidylserine cofactor, PS). Protein kinase C also phosphorylates enzymes that mediate specific actions associated with extrinsic receptor stimulation R . Branches of the chain of reactions involving IF h and DAG can be activated independently by ionomycin and phorbol ester, respectively (modified)
convert the reaction between a transmitter and an outer membrane receptor into phosphorylation of a variety of intracellular proteins, which can then influence various cell functions. One of the significant aspects of the problem is that, as far as is known today, there are only these two closely related regulatory systems of this type, used by numerous external mediators to regulate a variety of intracellular processes. At the same time, these regulatory systems, including Ca 2+, closely interact with each other, which allows them to carry out fine regulation of cellular functions.
Tutorials and Guides
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts TO., Watson J.D.Molecular Biology of the Cell, New York and London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biology. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille IN. Ionic channels of excitable membranes. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biophysics. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann TO., Mahler H.Biochemie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Principles of neural science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Original articles and reviews
8. Berridge M. J. The molecular basis of communication within the cell, Sci. Amer, 253, 124-134 (1985).
9. Berridge M. J., Irvine R. F. Inositol triphosphate, a novel second messenger in cellular signal transduction. Nature, 312, 315 321 (1984).
10. Bretscher M.S. The molecules of the cell membrane, Sci. Amer., 253, 124–134 (1985).
11. Daut J. The living cell as an energy–transducing machine. A minimal model of myocardial metabolism, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz IN. The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Active transport of cations in giant axons from Sepia and Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Longer P. Ionic channels with conformational substates, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Exoplasmic transport in normal and pathologic systems. In: Physiology and Pathology of Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
We invite you to familiarize yourself with the materials and.
: cellulose membrane, membrane, cytoplasm with organelles, nucleus, vacuoles with cell sap.The presence of plastids is the main feature of a plant cell.
Functions of the cell membrane- determines the shape of the cell, protects against environmental factors.
Plasma membrane- a thin film, consisting of interacting molecules of lipids and proteins, delimits the internal contents from the external environment, ensures the transport of water, minerals and organic substances into the cell by osmosis and active transport, and also removes waste products.
Cytoplasm- the internal semi-liquid environment of the cell, in which the nucleus and organelles are located, provides connections between them, and participates in basic life processes.
Endoplasmic reticulum- a network of branching channels in the cytoplasm. It is involved in the synthesis of proteins, lipids and carbohydrates, and in the transport of substances. Ribosomes are bodies located on the ER or in the cytoplasm, consisting of RNA and protein, and are involved in protein synthesis. EPS and ribosomes are a single apparatus for the synthesis and transport of proteins.
Mitochondria- organelles delimited from the cytoplasm by two membranes. Organic substances are oxidized in them and ATP molecules are synthesized with the participation of enzymes. Increase in the surface of the inner membrane on which enzymes are located due to cristae. ATP is an energy-rich organic substance.
Plastids(chloroplasts, leucoplasts, chromoplasts), their content in the cell is the main feature of the plant organism. Chloroplasts are plastids containing the green pigment chlorophyll, which absorbs light energy and uses it to synthesize organic substances from carbon dioxide and water. Chloroplasts are separated from the cytoplasm by two membranes, numerous outgrowths - grana on the inner membrane, in which chlorophyll molecules and enzymes are located.
Golgi complex- a system of cavities delimited from the cytoplasm by a membrane. The accumulation of proteins, fats and carbohydrates in them. Carrying out the synthesis of fats and carbohydrates on membranes.
Lysosomes- bodies delimited from the cytoplasm by a single membrane. The enzymes they contain accelerate the breakdown of complex molecules into simple ones: proteins into amino acids, complex carbohydrates into simple ones, lipids into glycerol and fatty acids, and also destroy dead parts of the cell and entire cells.
Vacuoles- cavities in the cytoplasm filled with cell sap, a place of accumulation of reserve nutrients and harmful substances; they regulate the water content in the cell.
Core- the main part of the cell, covered on the outside with a two-membrane, pore-pierced nuclear envelope. Substances enter the core and are removed from it through the pores. Chromosomes are carriers of hereditary information about the characteristics of an organism, the main structures of the nucleus, each of which consists of one DNA molecule combined with proteins. The nucleus is the site of DNA, mRNA, and rRNA synthesis.

The presence of an outer membrane, cytoplasm with organelles, and a nucleus with chromosomes.
Outer or plasma membrane- delimits the contents of the cell from the environment (other cells, intercellular substance), consists of lipid and protein molecules, ensures communication between cells, transport of substances into the cell (pinocytosis, phagocytosis) and out of the cell.
Cytoplasm- the internal semi-liquid environment of the cell, which provides communication between the nucleus and organelles located in it. The main life processes take place in the cytoplasm.
Cell organelles:
1) endoplasmic reticulum (ER)- a system of branching tubules, participates in the synthesis of proteins, lipids and carbohydrates, in the transport of substances in the cell;
2) ribosomes- bodies containing rRNA are located on the ER and in the cytoplasm and participate in protein synthesis. EPS and ribosomes are a single apparatus for protein synthesis and transport;
3) mitochondria- “power stations” of the cell, delimited from the cytoplasm by two membranes. The inner one forms cristae (folds), increasing its surface. Enzymes on the cristae accelerate the oxidation of organic substances and the synthesis of energy-rich ATP molecules;
4) Golgi complex- a group of cavities delimited by a membrane from the cytoplasm, filled with proteins, fats and carbohydrates, which are either used in vital processes or removed from the cell. The membranes of the complex carry out the synthesis of fats and carbohydrates;
5) lysosomes- bodies filled with enzymes accelerate the breakdown of proteins into amino acids, lipids into glycerol and fatty acids, polysaccharides into monosaccharides. In lysosomes, dead parts of the cell, whole cells, are destroyed.
Cellular inclusions- accumulations of reserve nutrients: proteins, fats and carbohydrates.
Core- the most important part of the cell. It is covered with a double-membrane shell with pores, through which some substances penetrate into the nucleus, and others enter the cytoplasm. Chromosomes are the main structures of the nucleus, carriers of hereditary information about the characteristics of the organism. It is transmitted during the division of the mother cell to daughter cells, and with germ cells to daughter organisms. The nucleus is the site of DNA, mRNA, and rRNA synthesis.
Exercise:
Explain why organelles are called specialized cell structures?
Answer: organelles are called specialized cell structures, since they perform strictly defined functions, hereditary information is stored in the nucleus, ATP is synthesized in mitochondria, photosynthesis occurs in chloroplasts, etc.
If you have questions about cytology, you can contact
The third stage of evolution is the appearance of the cell.
Molecules of proteins and nucleic acids (DNA and RNA) form a biological cell, the smallest unit of living things. Biological cells are the “building blocks” of all living organisms and contain all the material codes of development.
For a long time, scientists considered the cell structure to be extremely simple. The Soviet encyclopedic dictionary interprets the concept of a cell as follows: “A cell is an elementary living system, the basis of the structure and vital activity of all animals and plants.” It should be noted that the term “elementary” in no way means “simplest.” On the contrary, a cell is a unique fractal creation of God, striking in its complexity and at the same time exceptional coherence of the work of all its elements.
When we managed to look inside with the help of an electron microscope, it turned out that the structure of the simplest cell is as complex and incomprehensible as the Universe itself. Today it has already been established that “The cell is a special matter of the Universe, a special matter of the Cosmos.” One single cell contains information that can only be contained in several tens of thousands of volumes of the Great Soviet Encyclopedia. Those. a cell, among other things, is a huge “bioreservoir” of information.”
The author of the modern theory of molecular evolution, Manfred Eigen, writes: “In order for a protein molecule to be formed by chance, nature would have to make approximately 10,130 tests and spend on this a number of molecules that would be enough for 1027 Universes. If the protein was built intelligently, that is, "that the validity of each move could be checked by some kind of selection mechanism, then this required only about 2000 attempts. We come to a paradoxical conclusion: the program for constructing a "primitive living cell" is encoded somewhere at the level of elementary particles."
And how could it be otherwise? Each cell, possessing DNA, is endowed with consciousness, is aware of itself and other cells, and is in contact with the Universe, being, in fact, a part of it. And although the number and diversity of cells in the human body is amazing (about 70 trillion), they are all self-similar, just as all processes occurring in cells are self-similar. As German scientist Roland Glaser puts it, the design of biological cells is “very well thought out.” Well thought out by whom?
The answer is simple: proteins, nucleic acids, living cells and all biological systems are the product of the creative activity of an intelligent Creator.
What is interesting: at the atomic level there are no differences between the chemical composition of the organic and inorganic world. In other words, at the atomic level, a cell is created from the same elements as inanimate nature. Differences are found at the molecular level. In living bodies, along with inorganic substances and water, there are also proteins, carbohydrates, fats, nucleic acids, the enzyme ATP synthase and other low-molecular organic compounds.
To date, the cell has literally been disassembled into atoms for the purpose of study. However, it is never possible to create even one living cell, because to create a cell means to create a particle of the living Universe. Academician V.P. Kaznacheev believes that “a cell is a cosmoplanetary organism... Human cells are certain systems of ether-torsion biocolliders. In these biocolliders, processes unknown to us occur, the materialization of cosmic forms of flows, their cosmotransformation takes place, and due to this, particles are materialized.”
Water.
Almost 80% of the cell's mass is water. According to Doctor of Biological Sciences S. Zenin, water, due to its cluster structure, is an information matrix for controlling biochemical processes. In addition, it is water that is the primary “target” with which sound frequency vibrations interact. The order of cellular water is so high (close to the order of a crystal) that it is called a liquid crystal.
Squirrels.
Proteins play a huge role in biological life. A cell contains several thousand proteins unique to this type of cell (with the exception of stem cells). The ability to synthesize exactly one's own proteins is inherited from cell to cell and persists throughout life. During the life of a cell, proteins gradually change their structure and their function is disrupted. These spent proteins are removed from the cell and replaced with new ones, due to which the vital activity of the cell is maintained.
Let us note, first of all, the construction function of proteins, because they are the building material from which the membranes of cells and cellular organelles, the walls of blood vessels, tendons, cartilage, etc. are composed.
The signaling function of proteins is extremely interesting. It turns out that proteins can serve as signaling substances, transmitting signals between tissues, cells or organisms. The signaling function is performed by hormone proteins. Cells can interact with each other at a distance using signaling proteins transmitted through the intercellular substance.
Proteins also have a motor function. All types of movement that cells are capable of, such as muscle contraction, are performed by special contractile proteins. Proteins also perform a transport function. They are able to attach various substances and transfer them from one place in the cell to another. For example, the blood protein hemoglobin attaches oxygen and carries it to all tissues and organs of the body. In addition, proteins also have a protective function. When foreign proteins or cells are introduced into the body, it produces special proteins that bind and neutralize foreign cells and substances. And finally, the energy function of proteins is that with the complete breakdown of 1g of protein, energy is released in the amount of 17.6 kJ.
Cell structure.
The cell consists of three inextricably linked parts: the membrane, the cytoplasm and the nucleus, and the structure and function of the nucleus are different at different periods of the cell’s life. For the life of a cell includes two periods: division, which results in the formation of two daughter cells, and the period between divisions, which is called interphase.
The cell membrane directly interacts with the external environment and interacts with neighboring cells. It consists of an outer layer and a plasma membrane located underneath it. The surface layer of animal cells is called glycocalis. It communicates cells with the external environment and with all the substances surrounding it. Its thickness is less than 1 micron.
Cell structure
The cell membrane is a very important part of the cell. It holds all cellular components together and delineates the external and internal environments.
There is a constant exchange of substances between cells and the external environment. Water, various salts in the form of individual ions, and inorganic and organic molecules enter the cell from the external environment. Metabolic products, as well as substances synthesized in the cell: proteins, carbohydrates, hormones that are produced in the cells of various glands, are removed into the external environment through the membrane from the cell. Transport of substances is one of the main functions of the plasma membrane.
Cytoplasm- internal semi-liquid environment in which the main metabolic processes occur. Recent studies have shown that the cytoplasm is not some kind of solution, the components of which interact with each other through random collisions. It can be compared to jelly, which begins to “quiver” in response to external influences. This is how the cytoplasm perceives and transmits information.
The cytoplasm contains the nucleus and various organelles, which are united by it into one whole, which ensures their interaction and the activity of the cell as a single integrated system. The nucleus is located in the central part of the cytoplasm. The entire internal zone of the cytoplasm is filled with the endoplasmic reticulum, which is a cellular organelle: a system of tubules, vesicles and “cisterns” delimited by membranes. The endoplasmic reticulum is involved in metabolic processes, ensuring the transport of substances from the environment into the cytoplasm and between individual intracellular structures, but its main function is participation in protein synthesis, which occurs in ribosomes. - microscopic round bodies with a diameter of 15-20 nm. Synthesized proteins first accumulate in the channels and cavities of the endoplasmic reticulum and are then transported to organelles and cell sites where they are consumed.
In addition to proteins, the cytoplasm also contains mitochondria, small bodies 0.2-7 microns in size, which are called the “power stations” of cells. Redox reactions take place in mitochondria, providing cells with energy. The number of mitochondria in one cell ranges from a few to several thousand.
Core- the vital part of the cell, controls the synthesis of proteins and through them all physiological processes in the cell. In the nucleus of a non-dividing cell, a nuclear envelope, nuclear sap, nucleolus and chromosomes are distinguished. Through the nuclear envelope, a continuous exchange of substances occurs between the nucleus and the cytoplasm. Under the nuclear envelope is the nuclear sap (semi-liquid substance), which contains the nucleolus and chromosomes. The nucleolus is a dense round body, the dimensions of which can vary widely, from 1 to 10 μm or more. It consists mainly of ribonucleoproteins; participates in the formation of ribosomes. Usually there are 1-3 nucleoli in a cell, sometimes up to several hundred. The nucleolus contains RNA and protein.
With the appearance of the cell on Earth, Life arose!
To be continued...


