Осигурява връзката на клетката с околната среда. Връзката на организма с околната среда
Връзката на организма с околната среда от физикохимична гледна точка е отворена система, тоест система, в която протичат биохимични процеси. Първоначалните вещества идват от околната среда, а веществата, които също се образуват непрекъснато, се извеждат. Балансът между скоростта и концентрацията на продуктите на многопосочните реакции в организма е условен, въображаем, тъй като приемът и отстраняването на веществата не спират. Непрекъсната връзка с околната среда и ни позволява да разглеждаме живия организъм като отворена система.
Слънцето е източник на енергия за всички живи клетки. Растителните клетки улавят енергията на слънчевата светлина с помощта на хлорофил, използвайки я за реакции на асимилация по време на фотосинтезата. Клетките на животни, гъбички, бактерии използват слънчевата енергия индиректно, когато разделят органични вещества, синтезирани от земно растение.
Част от хранителните вещества на клетката се разграждат в процеса на клетъчното дишане, като по този начин се доставя енергията, необходима за различни видове клетъчна дейност. Този процес протича в органели, наречени митохондрии. Митохондрият се състои от две мембрани: външната, която отделя органелата от цитоплазмата, и вътрешната, която образува множество гънки. Основният продукт на дишането е АТФ. Той напуска митохондриите и се използва като енергиен източник за много химични реакции в цитоплазмата и клетъчната мембрана. Ако за осъществяването на клетъчното дишане е необходим кислород, тогава дишането се нарича аеробно дишане, но ако реакциите протичат при липса на кислород, тогава се говори за анаеробно дишане.
За всякакъв вид работа, извършена в клетката, енергията се използва в единична форма - под формата на енергия от фосфатните връзки на АТФ. АТФ е силно подвижно съединение. Образуването на АТФ става във вътрешната мембрана на митохондриите. АТФ се синтезира във всички клетки по време на дишането поради енергията на окисление на въглехидрати, мазнини и други органични вещества. В клетките на зелените растения основното количество АТФ се синтезира в хлоропластите поради слънчевата енергия. В тях по време на фотосинтезата се произвежда многократно повече АТФ, отколкото в митохондриите. АТФ се разлага с разрушаването на връзките фосфор-кислород и освобождаването на енергия. Това се случва под действието на ензима АТФаза в процеса на хидролиза на АТФ - добавяне на вода с елиминиране на молекула на фосфорната киселина. В резултат на това АТФ се превръща в ADP и ако две молекули фосфорна киселина се отделят, след това в AMP. Реакцията на разцепване на всяка грам-молекула киселина е придружена от освобождаване на 40 kJ. Това е много голям добив на енергия, така че връзките фосфор-кислород на АТФ обикновено се наричат макроергични (високоенергийни).
Използването на АТФ в реакциите на пластичен обмен се осъществява чрез тяхното конюгиране с хидролизата на АТФ. Молекулите на различни вещества се зареждат с енергия чрез свързване на фосфорната група, освободена по време на хидролизата от молекулата на АТФ, тоест чрез фосфорилиране.
Характерна особеност на фосфатните производни е, че те не могат да напуснат клетката, въпреки че техните "разредени" форми свободно преминават през мембраната. Поради това фосфорилираните молекули остават в клетката, докато не бъдат използвани в съответните реакции.
Обратният процес на преобразуване на ADP в ATP се осъществява чрез прикрепване на молекула фосфорна киселина към ADP, освобождавайки вода и поглъщайки голямо количество енергия.
По този начин АТФ е универсален и непосредствен източник на енергия за клетъчната активност. Това създава единен клетъчен енергиен фонд и прави възможно преразпределянето и транспортирането му от една част на клетката в друга.
Прехвърлянето на фосфатна група играе важна роля в химичните реакции като сглобяването на макромолекули от мономери. Например, аминокиселините могат да бъдат комбинирани в пептиди само ако са били предварително фосфорилирани. Механичните процеси на свиване или движение, пренос на разтворено вещество срещу градиент на концентрация и други процеси са свързани с разхода на енергия, съхранявана в АТФ.
Процесът на обмен на енергия може да бъде представен по следния начин. Високомолекулните органични вещества в цитоплазмата ензимно, чрез хидролиза, се превръщат в по-прости, от които се състоят: протеини - в аминокиселини, поли- и дизахариди - в монозахариди (+ глюкоза), мазнини в глицерол и мастни киселини. Окислителните процеси отсъстват, отделя се малко енергия, която не се използва и преминава в термична форма. Повечето клетки използват първо въглехидратите. Полизахаридите (нишесте в растенията и гликоген в животните) се хидролизират до глюкоза. Окислението на глюкозата протича в три фази: гликолиза, окислително декарбоксилиране (цикъл на Кребс - цикъл на лимонена киселина) и окислително фосфорилиране (дихателна верига). В цитоплазмата се извършва гликолиза, в резултат на която една молекула глюкоза се разделя на две молекули пирогроздна киселина с освобождаване на две молекули АТФ. При липса на кислород, пирогрозената киселина се превръща или в етанол (ферментация), или в млечна киселина (анаеробно дишане).
Ако гликолизата се извършва в животински клетки, глюкозната молекула с шест въглерода се разпада на две молекули млечна киселина. Този процес е многоетапен. Извършва се последователно от 13 ензима. По време на алкохолната ферментация от молекула глюкоза се образуват две молекули етанол и две молекули CO2.
Гликолизата е фаза, обща за анаеробното и аеробното дишане, другите две се извършват само при аеробни условия. Процесът на безкислородно окисление, при който се освобождава и използва само част от енергията на метаболитите, е крайният процес за анаеробните организми. В присъствието на кислород пирогрозената киселина преминава в митохондриите, където в резултат на редица последователни реакции се окислява напълно аеробно до H2O и CO2 с едновременно фосфорилиране на ADP до ATP. В същото време гликолизата дава две молекули АТФ, две - цикъла на Кребс, 34 - дихателната верига. Нетният добив от пълното окисление на една молекула глюкоза до H2O и CO2 е 38 молекули.
По този начин при аеробните организми окончателното разлагане на органичните вещества се извършва чрез окисляването им с атмосферен кислород до прости неорганични вещества: CO2 и H2O. Този процес протича върху кристите на митохондриите. В този случай се освобождава максимално количество свободна енергия, значителна част от която е запазена в молекулите на АТФ. Лесно е да се види, че аеробното окисление осигурява на клетката безплатна енергия в най-голяма степен.
В резултат на катаболизма богатите на енергия АТФ молекули се натрупват в клетката, а CO2 и излишната вода се отделят във външната среда.
Захарните молекули, които не са необходими за дишането, могат да се съхраняват в клетката. Излишните липиди или се разцепват, след което продуктите от тяхното разцепване влизат в митохондриите като субстрат за дишане, или се отлагат в резерв в цитоплазмата под формата на мастни капчици. Протеините се изграждат от аминокиселини, влизащи в клетката. Синтезът на протеин се осъществява в органели, наречени рибозоми. Всяка рибозома се състои от две субчастици - голяма и малка: и двете субчастици включват протеинови молекули и РНК молекули.
Рибозомите често са прикрепени към специална система от мембрани, състояща се от резервоари и везикули, към така наречения ендоплазмен ретикулум (ER); в клетки, които произвеждат много протеин, ендоплазменият ретикулум често е много добре развит и е пълен с рибозоми. Някои ензими са ефективни само ако са прикрепени към мембрана. Повечето от ензимите, участващи в синтеза на липиди, се намират тук. По този начин ендоплазменият ретикулум е като че ли един вид клетъчен работен плот.
В допълнение, ER разделя цитоплазмата на отделни секции или отделения, т.е. разделя различните химични процеси, протичащи едновременно в цитоплазмата, и по този начин намалява вероятността тези процеси да се намесват един в друг.
Често продуктите, образувани от дадена клетка, се използват извън клетката. В такива случаи протеините, синтезирани върху рибозоми, преминават през мембраните на ендоплазмения ретикулум и се опаковат в мембранни везикули, които се образуват около тях, които след това се отделят от ER. Тези мехурчета, сплескващи се и подредени един върху друг, като палачинки в купчина, образуват характерна структура, наречена комплекс на Голджи или апарат на Голджи. По време на престоя си в апарата на Голджи протеините претърпяват определени промени. Когато дойде време да напуснат клетката, мембранозните везикули се сливат с клетъчната мембрана и се изпразват, изливайки съдържанието си, т.е. секрецията се осъществява чрез екзоцитоза.
Лизозомите се образуват и в апарата на Голджи – мембранни торбички, съдържащи храносмилателни ензими. Разбирането как една клетка произвежда, пакетира и изнася определени протеини и как „знае“ кои протеини трябва да запази за себе си, е един от най-завладяващите клонове на съвременната цитология.
Мембраните на всяка клетка непрекъснато се движат и променят. ER мембраните се движат бавно в клетката. Отделни участъци от тези мембрани се отделят и образуват везикули, които за известно време стават част от апарата на Голджи, а след това, в процеса на екзоцитоза, се сливат с клетъчната мембрана.
По-късно мембранният материал се връща в цитоплазмата, където се използва повторно.
Обмяната на веществата, влизащи в клетката или освободени от нея навън, както и обменът на различни сигнали с микро- и макросредата, става през външната мембрана на клетката. Както е известно, клетъчната мембрана е липиден бислой, в който са вградени различни протеинови молекули, които действат като специализирани рецептори, йонни канали, устройства, които активно пренасят или отстраняват различни химикали, междуклетъчни контакти и т.н. В здравите еукариотни клетки фосфолипидите се разпределят в мембраната асиметрична: външната повърхност се състои от сфингомиелин и фосфатидилхолин, вътрешната повърхност се състои от фосфатидилсерин и фосфатидилетаноламин. Поддържането на такава асиметрия изисква енергийни разходи. Следователно, при увреждане на клетката, нейната инфекция, енергиен глад, външната повърхност на мембраната се обогатява с необичайни за нея фосфолипиди, което се превръща в сигнал за други клетки и ензими за клетъчно увреждане с подходяща реакция към него. Най-важна роля играе разтворимата форма на фосфолипаза А2, която разгражда арахидоновата киселина и създава лизоформи от горните фосфолипиди. Арахидоновата киселина е ограничаващо звено за създаването на такива възпалителни медиатори като ейкозаноидите, а защитните молекули - пентраксини (С-реактивен протеин (CRP), прекурсори на амилоидните протеини) - се прикрепват към лизоформите в мембраната, последвано от активиране на система на комплемента по класическия път и клетъчно разрушаване.
Структурата на мембраната допринася за запазването на характеристиките на вътрешната среда на клетката, нейните разлики от външната среда. Това се осигурява от селективната пропускливост на клетъчната мембрана, наличието на активни транспортни механизми в нея. Нарушаването им в резултат на директно увреждане, например от тетродотоксин, уабаин, тетраетиламоний, или в случай на недостатъчно енергийно снабдяване на съответните „помпи“, води до нарушаване на електролитния състав на клетката, промяна в нейния метаболизъм , нарушение на специфични функции - свиване, провеждане на импулс на възбуждане и др. Нарушаването на клетъчните йонни канали (калций, натрий, калий и хлорид) при хората може да бъде генетично обусловено и от мутация на гените, отговорни за структурата на тези канали. Така наречените каналопатии са причина за наследствени заболявания на нервната, мускулната и храносмилателната система. Прекомерният прием на вода вътре в клетката може да доведе до нейното разкъсване - цитолиза - поради перфорация на мембраната по време на активиране на комплемента или атака от цитотоксични лимфоцити и естествени убийци.
В клетъчната мембрана са вградени много рецептори – структури, които, когато се комбинират със съответните специфични сигнални молекули (лиганди), предават сигнал в клетката. Това се случва чрез различни регулаторни каскади, състоящи се от ензимно активни молекули, които се активират последователно и в крайна сметка допринасят за изпълнението на различни клетъчни програми, като растеж и пролиферация, диференциация, подвижност, стареене и клетъчна смърт. Регулаторните каскади са доста многобройни, но броят им все още не е напълно определен. Системата от рецептори и свързаните с тях регулаторни каскади също съществуват вътре в клетката; те създават определена регулаторна мрежа с точки на концентрация, разпределение и избор на по-нататъшен сигнален път в зависимост от функционалното състояние на клетката, етапа на нейното развитие и едновременното действие на сигнали от други рецептори. Резултатът от това може да бъде инхибиране или усилване на сигнала, неговата посока по различен регулаторен път. Както рецепторният апарат, така и пътищата на сигнална трансдукция през регулаторни каскади, например към ядрото, могат да бъдат нарушени в резултат на генетичен дефект, който възниква като вроден дефект на нивото на организма или поради соматична мутация в определена клетка Тип. Тези механизми могат да бъдат повредени от инфекциозни агенти, токсини, а също и да се променят по време на стареенето. Последният етап от това може да бъде нарушение на функциите на клетката, процесите на нейната пролиферация и диференциация.
На повърхността на клетките се намират и молекули, които играят важна роля в процесите на междуклетъчно взаимодействие. Те могат да включват протеини на клетъчна адхезия, антигени за тъканна съвместимост, тъканно-специфични, диференциращи антигени и др. Промените в състава на тези молекули причиняват нарушение на междуклетъчните взаимодействия и могат да предизвикат активиране на съответните механизми за елиминиране на такива клетки, т.к. те представляват известна опасност за целостта на организма като резервоар на инфекция, особено вирусна, или като потенциални инициатори на туморен растеж.
Нарушаване на енергийното снабдяване на клетката
Източникът на енергия в клетката е храната, след разграждането на която енергията се освобождава до крайни вещества. Митохондриите са основното място за производство на енергия, в което веществата се окисляват с помощта на ензими на дихателната верига. Окислението е основният доставчик на енергия, тъй като в резултат на гликолизата не се освобождава повече от 5% от енергията от същото количество окислителни субстрати (глюкоза), в сравнение с окислението. Около 60% от енергията, освободена при окисляване, се натрупва чрез окислително фосфорилиране в макроергични фосфати (АТФ, креатин фосфат), останалата част се разсейва като топлина. В бъдеще високоенергийните фосфати се използват от клетката за процеси като изпомпване, синтез, делене, движение, секреция и т.н. Има три механизма, чието увреждане може да доведе до нарушаване на доставката на енергия към клетката : първият е механизмът за синтез на ензими на енергийния метаболизъм, вторият е механизмът на окислително фосфорилиране, третият - механизмът на използване на енергията.
Нарушаването на транспорта на електрони в дихателната верига на митохондриите или разединяването на окислението и фосфорилирането на ADP със загуба на протонния потенциал - движещата сила на генерирането на АТФ, води до отслабване на окислителното фосфорилиране по такъв начин, че по-голямата част от енергията се разсейва в формата на топлина и броят на макроергичните съединения намалява. Разединяването на окислението и фосфорилирането под въздействието на адреналина се използва от клетките на хомойотермните организми за увеличаване на производството на топлина, като същевременно се поддържа постоянна телесна температура по време на охлаждане или нейното повишаване по време на треска. При тиреотоксикоза се наблюдават значителни промени в структурата на митохондриите и енергийния метаболизъм. Тези промени първоначално са обратими, но след определен момент стават необратими: митохондриите се фрагментират, разпадат се или набъбват, губят кристи, превръщайки се във вакуоли и в крайна сметка натрупват вещества като хиалин, феритин, калций, липофусцин. При пациенти със скорбут, митохондриите се сливат, за да образуват хондриосфери, вероятно поради увреждане на мембраната от пероксидни съединения. Значително увреждане на митохондриите възниква под въздействието на йонизиращо лъчение, по време на трансформацията на нормална клетка в злокачествена.
Митохондриите са мощно депо от калциеви йони, където концентрацията му е с няколко порядъка по-висока от тази в цитоплазмата. Когато митохондриите са увредени, калцият навлиза в цитоплазмата, причинявайки активиране на протеинази с увреждане на вътреклетъчните структури и нарушаване на функциите на съответната клетка, например, калциеви контрактури или дори „калциева смърт“ в невроните. В резултат на нарушение на функционалната способност на митохондриите рязко се увеличава образуването на свободни радикални пероксидни съединения, които имат много висока реактивност и поради това увреждат важни клетъчни компоненти - нуклеинови киселини, протеини и липиди. Това явление се наблюдава по време на така наречения оксидативен стрес и може да има негативни последици за съществуването на клетката. По този начин увреждането на външната митохондриална мембрана е придружено от освобождаване в цитоплазмата на вещества, съдържащи се в междумембранното пространство, предимно цитохром С и някои други биологично активни вещества, които предизвикват верижни реакции, които причиняват програмирана клетъчна смърт - апоптоза. Увреждайки митохондриалната ДНК, реакциите на свободните радикали изкривяват генетичната информация, необходима за образуването на определени ензими на дихателната верига, които се произвеждат специално в митохондриите. Това води до още по-голямо нарушаване на окислителните процеси. Като цяло, присъщият генетичен апарат на митохондриите, в сравнение с генетичния апарат на ядрото, е по-малко защитен от вредни влияния, способни да променят генетичната информация, кодирана в него. В резултат на това митохондриалната дисфункция възниква през целия живот, например в процеса на стареене, по време на злокачествена трансформация на клетката, както и на фона на наследствени митохондриални заболявания, свързани с мутацията на митохондриалната ДНК в яйцеклетката. В момента са описани повече от 50 митохондриални мутации, които причиняват наследствени дегенеративни заболявания на нервната и мускулната система. Те се предават на детето изключително от майката, тъй като митохондриите на сперматозоидите не са част от зиготата и съответно новия организъм.
Нарушаване на съхранението и предаването на генетична информация
Клетъчното ядро съдържа по-голямата част от генетичната информация и по този начин осигурява нормалното му функциониране. С помощта на селективна генна експресия координира работата на клетката в интерфаза, съхранява генетична информация, пресъздава и пренася генетичен материал в процеса на клетъчно делене. Репликацията на ДНК и транскрипцията на РНК се извършват в ядрото. Различни патогенни фактори, като ултравиолетова и йонизираща радиация, окисление на свободните радикали, химикали, вируси, могат да увредят ДНК. Смята се, че всяка клетка на топлокръвно животно за 1 ден. губи над 10 000 бази. Към това трябва да се добавят и нарушения при копиране по време на разделяне. Ако това увреждане продължи, клетката не би могла да оцелее. Защитата се състои в съществуването на мощни системи за възстановяване, като ултравиолетова ендонуклеаза, репаративна репликация и рекомбинационни възстановителни системи, които заместват увреждането на ДНК. Генетичните дефекти в репаративните системи предизвикват развитие на заболявания поради повишена чувствителност към фактори, които увреждат ДНК. Това е пигментна ксеродермия, както и някои синдроми на ускорено стареене, придружени от повишена склонност към поява на злокачествени тумори.
Системата за регулиране на процесите на репликация на ДНК, транскрипция на информационна РНК (иРНК), транслация на генетична информация от нуклеинови киселини в структурата на протеините е доста сложна и многостепенна. В допълнение към регулаторните каскади, които задействат действието на повече от 3000 транскрипционни фактора, които активират определени гени, съществува и многостепенна регулаторна система, медиирана от малки РНК молекули (интерфериращи РНК; RNAi). Човешкият геном, който се състои от приблизително 3 милиарда пуринови и пиримидинови бази, съдържа само 2% от структурните гени, отговорни за протеиновия синтез. Останалите осигуряват синтеза на регулаторни РНК, които заедно с транскрипционните фактори активират или блокират работата на структурните гени на ниво ДНК в хромозомите или влияят на транслацията на информационната РНК (иРНК) по време на образуването на полипептидна молекула в цитоплазмата . Нарушаването на генетичната информация може да се случи както на ниво структурни гени, така и на регулаторната част на ДНК със съответните прояви под формата на различни наследствени заболявания.
Напоследък се отделя голямо внимание на промените в генетичния материал, които настъпват по време на индивидуалното развитие на организма и са свързани с инхибирането или активирането на определени участъци от ДНК и хромозоми поради тяхното метилиране, ацетилиране и фосфорилиране. Тези промени продължават дълго време, понякога през целия живот на организма от ембриогенезата до старостта и се наричат епигеномно наследяване.
Възпроизвеждането на клетки с променена генетична информация също се предотвратява от системите (фактори) за контрол на митотичния цикъл. Те взаимодействат с циклин-зависимите протеин кинази и техните каталитични субединици - циклини - и блокират преминаването на пълен митотичен цикъл от клетката, спирайки деленето на границата между предсинтетичната и синтетичната фаза (блок G1 / S), докато възстановяването на ДНК завърши , а ако е невъзможно, те инициират програмирани клетки на смърт. Тези фактори включват гена p53, чиято мутация причинява загуба на контрол върху пролиферацията на трансформирани клетки; среща се при почти 50% от раковите заболявания при хората. Вторият контролно-пропускателен пункт на преминаването на митотичния цикъл се намира на границата G2/M. Тук правилното разпределение на хромозомния материал между дъщерните клетки в митоза или мейоза се контролира с помощта на комплекс от механизми, които контролират клетъчното вретено, центъра и центромерите (кинетохори). Неефективността на тези механизми води до нарушаване на разпределението на хромозомите или техните части, което се проявява чрез липса на каквато и да е хромозома в една от дъщерните клетки (анеуплоидия), наличие на допълнителна хромозома (полиплоидия), отделяне на част от хромозомата (делеция) и нейното прехвърляне към друга хромозома (транслокация). Такива процеси много често се наблюдават при размножаването на злокачествено изродени и трансформирани клетки. Ако това се случи по време на мейоза със зародишни клетки, това води или до смъртта на плода в ранен стадий на ембрионално развитие, или до раждането на организъм с хромозомно заболяване.
Неконтролираното размножаване на клетките по време на растежа на тумора възниква в резултат на мутации в гените, които контролират клетъчната пролиферация и се наричат онкогени. Сред над 70-те известни в момента онкогени, повечето от тях са компоненти на регулацията на клетъчния растеж, някои са транскрипционни фактори, които регулират генната активност, както и фактори, които инхибират клетъчното делене и растеж. Друг фактор, ограничаващ прекомерното разширяване (разпространение) на пролифериращи клетки, е скъсяването на краищата на хромозомите - теломерите, които не могат да се репликират напълно в резултат на чисто стерично взаимодействие, следователно след всяко клетъчно делене теломерите се съкращават с определена част от основите. Така пролифериращите клетки на възрастен организъм след определен брой деления (обикновено от 20 до 100, в зависимост от вида на организма и неговата възраст) изчерпват дължината на теломерите и по-нататъшната репликация на хромозомите спира. Това явление не се среща в сперматогенния епител, ентероцитите и ембрионалните клетки поради наличието на ензима теломераза, който възстановява дължината на теломерите след всяко делене. В повечето клетки на възрастни организми теломеразата е блокирана, но, за съжаление, тя се активира в туморните клетки.
Връзката между ядрото и цитоплазмата, транспортирането на вещества в двете посоки се осъществяват през порите в ядрената мембрана с участието на специални транспортни системи с консумация на енергия. Така енергийни и пластични вещества, сигнални молекули (транскрипционни фактори) се транспортират до ядрото. Обратният поток въвежда в цитоплазмата молекули на иРНК и трансферна РНК (тРНК), рибозоми, необходими за протеиновия синтез в клетката. Същият начин на транспортиране на вещества е присъщ на вирусите, по-специално като ХИВ. Те пренасят своя генетичен материал в ядрото на клетката гостоприемник с по-нататъшното му включване в генома на гостоприемника и пренасянето на новообразуваната вирусна РНК в цитоплазмата за по-нататъшен протеинов синтез на нови вирусни частици.
Нарушаване на процесите на синтез
Процесите на протеинов синтез протичат в цистерните на ендоплазмения ретикулум, тясно свързани с порите в ядрената мембрана, през които рибозомите, тРНК и иРНК навлизат в ендоплазмения ретикулум. Тук се осъществява синтеза на полипептидни вериги, които по-късно придобиват окончателната си форма в агрануларния ендоплазмен ретикулум и ламеларния комплекс (комплекс на Голджи), където претърпяват посттранслационна модификация и свързване с въглехидратни и липидни молекули. Новообразуваните протеинови молекули не остават на мястото на синтеза, а с помощта на сложен регулиран процес, който се нарича протеин кинеза, се пренасят активно в онази изолирана част на клетката, където ще изпълняват предназначената си функция. В този случай много важна стъпка е структурирането на прехвърлената молекула в подходяща пространствена конфигурация, способна да изпълнява присъщата си функция. Такова структуриране става с помощта на специални ензими или върху матрица от специализирани протеинови молекули - шаперони, които помагат на новообразуваната или изменена поради външно въздействие протеинова молекула да придобие правилната триизмерна структура. В случай на неблагоприятен ефект върху клетката, когато има възможност за нарушаване на структурата на протеиновите молекули (например с повишаване на телесната температура, инфекциозен процес, интоксикация), концентрацията на шаперони в клетката нараства рязко. Следователно такива молекули също се наричат стрес протеини, или протеини от топлинен шок. Нарушаването на структурирането на протеиновата молекула води до образуването на химически инертни конгломерати, които се отлагат в клетката или извън нея в случай на амилоидоза, болест на Алцхаймер и др. Понякога предварително структурирана аналогична молекула може да служи като матрица и в в този случай, ако първичното структуриране не е настъпило правилно, всички следващи молекули също ще бъдат дефектни. Тази ситуация възниква при така наречените прионни заболявания (скрапи при овце, бяс при крави, куру, болест на Кройцфелд-Якоб при хората), когато дефект в един от мембранните протеини на нервната клетка причинява последващо натрупване на инертни маси вътре клетката и нарушаване на нейната жизнена дейност.
Нарушаването на процесите на синтез в клетката може да настъпи на различни етапи: транскрипция на РНК в ядрото, транслация на полипептиди в рибозоми, посттранслационна модификация, хиперметилиране и гликозилиране на бежовата молекула, транспорт и разпределение на протеини в клетката и тяхното отстраняване навън. В този случай може да се наблюдава увеличаване или намаляване на броя на рибозомите, разпадане на полирибозомите, разширяване на цистерните на гранулирания ендоплазмен ретикулум, загуба на рибозоми от него, образуване на везикули и вакуоли. Така че, в случай на отравяне с бледа гъба, ензимът РНК полимераза се уврежда, което нарушава транскрипцията. Дифтерийният токсин, инактивирайки фактора на удължаване, нарушава процесите на транслация, причинявайки увреждане на миокарда. Причината за нарушаване на синтеза на някои специфични протеинови молекули могат да бъдат инфекциозни агенти. Например, херпесвирусите инхибират синтеза и експресията на МНС антигенни молекули, което им позволява частично да избягват имунния контрол, а чумните бацили инхибират синтеза на медиатори на остро възпаление. Появата на необичайни протеини може да спре по-нататъшното им разграждане и да доведе до натрупване на инертен или дори токсичен материал. До известна степен за това може да допринесе и нарушаването на процесите на разпад.
Нарушаване на процесите на гниене
Едновременно със синтеза на белтъка в клетката непрекъснато протича разпадането му. При нормални условия това има важно регулаторно и формиращо значение, например по време на активирането на неактивни форми на ензими, протеинови хормони и протеини от митотичния цикъл. Нормалният клетъчен растеж и развитие изискват фино контролиран баланс между синтеза и разграждането на протеини и органели. Въпреки това, в процеса на протеинов синтез, поради грешки в работата на синтезиращия апарат, ненормално структуриране на протеиновата молекула, нейното увреждане от химически и бактериални агенти, постоянно се образуват доста голям брой дефектни молекули. Според някои оценки техният дял е около една трета от всички синтезирани протеини.
Клетките на бозайниците имат няколко основни пътища на разграждане на протеини:чрез лизозомни протеази (пентид хидролази), калций-зависими протеинази (ендопептидази) и протеазомната система. Освен това има и специализирани протеинази, като каспази. Основната органела, в която се извършва разграждането на веществата в еукариотните клетки, е лизозомата, която съдържа множество хидролитични ензими. Поради процесите на ендоцитоза и различни видове аутофагия в лизозомите и фаголизозомите се унищожават както дефектни протеинови молекули, така и цели органели: увредени митохондрии, участъци от плазмената мембрана, някои извънклетъчни протеини, съдържанието на секреторни гранули.
Важен механизъм за разграждане на протеина е протеазомата, сложна мултикаталитична протеиназна структура, локализирана в цитозола, ядрото, ендоплазмения ретикулум и върху клетъчната мембрана. Тази ензимна система е отговорна за разграждането на увредените протеини, както и здравите протеини, които трябва да бъдат отстранени за нормална клетъчна функция. В този случай протеините, които трябва да бъдат унищожени, предварително се комбинират със специфичен убиквитин полипептид. Въпреки това, протеини, които не са повсеместни, също могат да бъдат частично унищожени в протеазомите. Разграждането на протеинова молекула в протеазоми до къси полипептиди (обработка) с последващото им представяне заедно с МНС тип I молекули е важна връзка в осъществяването на имунния контрол на антигенната хомеостаза на организма. При отслабване на функцията на протеазомата се получава натрупване на увредени и ненужни протеини, което съпътства стареенето на клетките. Нарушаването на разграждането на циклин-зависимите протеини води до нарушение на клетъчното делене, разграждането на секреторните протеини - до развитие на цистофиброза. Обратно, увеличаването на протеазомната функция съпътства изчерпването на тялото (СПИН, рак).
При генетично обусловени нарушения на белтъчната деградация, организмът не е жизнеспособен и умира в ранните етапи на ембриогенезата. Ако разграждането на мазнините или въглехидратите е нарушено, тогава възникват заболявания на натрупване (тезавризми). В същото време вътре в клетката се натрупва прекомерно количество от определени вещества или продукти от непълното им разпадане – липиди, полизахариди, което значително уврежда функцията на клетката. Най-често се наблюдава в чернодробни епителиоцити (хепатоцити), неврони, фибробласти и макрофагоцити.
Придобитите нарушения в процесите на разпадане на веществата могат да възникнат в резултат на патологични процеси (например протеинова, мастна, въглехидратна и пигментна дистрофия) и да бъдат придружени от образуване на необичайни вещества. Нарушенията в системата на лизозомната протеолиза водят до намаляване на адаптацията по време на гладуване или повишено натоварване, до появата на някои ендокринни дисфункции - намаляване на нивото на инсулин, тиреоглобулин, цитокини и техните рецептори. Нарушенията на разграждането на протеините забавят скоростта на заздравяване на рани, причиняват развитие на атеросклероза и засягат имунния отговор. При хипоксия, промени във вътреклетъчното pH, радиационно увреждане, характеризиращо се с повишена пероксидация на мембранните липиди, както и под въздействието на лизозомотропни вещества - бактериални ендотоксини, метаболити на токсични гъби (спорофусарин), кристали от силициев оксид - стабилност на лизозомната мембрана промени, активирани лизозомни ензими се освобождават в цитоплазмата, което причинява разрушаване на клетъчните структури и нейната смърт.
Глава 1
ОСНОВИ НА КЛЕТЪЧНАТА ФИЗИОЛОГИЯ
И. Дюдел
плазмената мембрана . Животинските клетки са ограничени от плазмената мембрана (фиг. 1.1). Ще се спрем по-подробно на неговата структура, която е много подобна на структурата на много вътреклетъчни мембрани. Основната матрица на мембраната се състои от липидиглавно фосфатидилхолин. Тези липиди се състоят от хидрофилна главна група, към която са прикрепени дълги хидрофобни въглеводородни вериги. Във вода такива липиди спонтанно образуват двуслоен филм с дебелина 4-5 nm, в който хидрофилните групи са обърнати към водната среда, а хидрофобните въглеводородни вериги са подредени в два реда, образувайки безводна липидна фаза. Клетъчните мембрани са липидни двуслоеве от този тип и съдържат гликолипиди, холестерол и фосфолипиди (фиг. 1.2). Хидрофилната част на гликолипидите се образува от олигозахариди. Гликолипидите винаги са разположени на външната повърхност на плазмената мембрана, а олигозахаридната част на молекулата е ориентирана като косъм, потопен в околната среда. Разпръснати сред фосфолипидите в почти равни количества холестеролни молекули стабилизират мембраната. Разпределението на различните липиди във вътрешния и външния слой на мембраната не е еднакво и дори в рамките на един и същи слой има области, в които са концентрирани определени видове липиди. Такова неравномерно разпределение
Ориз. 1.1. Схематичен чертеж на клетка, показващ най-важните органели
вероятно има някакво, все още неясно, функционално значение.
Основните функционални елементи, потопени в относително инертния липиден матрикс на мембраната са катерици(фиг. 1.2). Протеинът по тегло варира от 25 до 75% в различни мембрани, но тъй като протеиновите молекули са много по-големи от липидните молекули, 50% тегловни са еквивалентни на съотношение от 1 протеинова молекула към 50 липидни молекули. Някои протеини проникват през мембраната от външната й към вътрешната повърхност, докато други са фиксирани в един слой. Протеиновите молекули обикновено са ориентирани така, че техните хидрофобни групи са потопени в липидната мембрана, а полярните хидрофилни групи на повърхността на мембраната са потопени във водната фаза. Много протеини от външната повърхност на мембраната са гликопротеини; техните хидрофилни захаридни групи са изправени пред извънклетъчната среда.
Мембранни системи на вътреклетъчните органели .
Приблизително половината от клетъчния обем се заема от органели, изолирани от цитозола чрез мембрани. Общата повърхност на мембраните на вътреклетъчните органели е поне 10 пъти по-голяма от повърхността на плазмената мембрана. Най-широко използваната мембранна система е ендоплазмения ретикулум,представляваща мрежа

Ориз. 1.2.Схематично представяне на плазмената мембрана. Протеините са вградени във фосфолипидния двуслой, като някои обхващат двуслоя, докато други са закотвени само към външния или вътрешния слой
силно извити тубули или сакуларни удължени структури; големи участъци от ендоплазмения ретикулум са осеяни с рибозоми; такъв ретикулум се нарича гранулиран, или груб (фиг. 1.1). апарат на Голджисъщо се състои от свързани с мембрана ламели, от които се откъсват везикули или везикули (фиг. 1.1). Лизозоми и пероксизомиса малки специализирани везикули. Във всички тези разнообразни органели, мембраната и пространството, което покрива, съдържат специфични набори от ензими; вътре в органелите се натрупват специални метаболитни продукти, които се използват за осъществяване на различни функции на органелите.
ЯдроИ митохондриисе различават по това, че всяка от тези органели е заобиколена от две мембрани. Ядрото е отговорно за кинетичния контрол на метаболизма; нагънатата вътрешна митохондриална мембрана е мястото на окислителния метаболизъм; тук, поради окисляването на пируват или мастни киселини, се синтезира високоенергийно съединение аденозин трифосфат (АТФ или АТФ).
цитоскелет . Цитоплазмата около органелите в никакъв случай не може да се счита за аморфна; тя е пронизана от мрежа от цитоскелет. Цитоскелетът се състои от микротубули, актинови филаменти и междинни филаменти (фиг. 1.1). микротубулиимат външен диаметър от около 25 nm; те се образуват, като нормален полимер, в резултат на сглобяването на тубулинови протеинови молекули. актинови нишки -контрактилни влакна, разположени в близкия мембранен слой и в цялата клетка - участват главно в процесите, свързани с движението. Междинни нишкисъстоят се от блокове с различен химичен състав в различни видове клетки; те образуват различни връзки между другите два елемента на цитоскелета, споменати по-горе. Органелите и плазмената мембрана също са свързани с цитоскелета, който не само поддържа формата на клетката и положението на органелите в нея, но и определя промяната във формата на клетката и нейната подвижност.
Цитозол . Около половината от обема на клетката се заема от цитозола. Тъй като е приблизително 20% (тегловно) протеин, той е по-скоро гел, отколкото воден разтвор. Малки молекули, включително органични и неорганични йони,разтворен във водната фаза. Между клетката и околната среда (извънклетъчното пространство) се осъществява обмен на йони; тези обменни процеси ще бъдат обсъдени в следващия раздел. Концентрацията на йони в извънклетъчното пространство се поддържа със значителна точност на постоянно ниво; вътреклетъчната концентрация на всеки от йоните също има специфично ниво, което се различава от това извън клетката (Таблица 1.1). Най-често срещаният катион в извънклетъчната среда е Na+ в клетката концентрацията му е повече от 10 пъти по-ниска. Напротив, вътре в клетката концентрацията на К + е най-висока, извън клетката е по-ниска с повече от порядък. Най-големият градиент между извънклетъчните и вътреклетъчните концентрации съществува за Ca 2+, чиято концентрация на свободни йони вътре в клетката е поне 10 000 пъти по-ниска от тази извън нея. Не всички йони се разтварят в цитозола; някои от тях се адсорбират върху протеини или се отлагат в органели. Например, в случая на Ca 2+ свързаните йони са много по-многобройни от свободните. Повечето от цитозолните протеини са ензими, с участието на които се осъществяват много процеси на междинен метаболизъм: гликолиза и глюконеогенеза, синтез или разрушаване на аминокиселини, протеинов синтез върху рибозоми (фиг. 1.1). Цитозолът съдържа също липидни капчици и гликогенови гранули, които служат като резерви от важни молекули.
Таблица 1.1.Вътре- и извънклетъчни концентрации на йони в мускулни клетки на хомойотермични животни. НО – – „клетъчни аниони с високо молекулно тегло“
|
Вътреклетъчна концентрация |
Извънклетъчна концентрация |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
С л - |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO 3 - |
8 mmol/l |
HCO 3 - |
27 mmol/l |
|
А- |
155 mmol/l |
Друго катиони |
5 mmol/l |
|
Потенциал на покой -90 mV |
|||
1.2. Обменът на вещества между клетката и околната среда
Описахме накратко структурата на клетката, за да използваме това описание за преглед на основите на клетъчната физиология. В никакъв случай клетката не може да се счита за статична формация, тъй като има постоянен обмен на вещества между различни вътреклетъчни отделения, както и между отделения и околната среда. Структурите на клетката са в динамично равновесие, а взаимодействието на клетките помежду си и с външната среда е необходимо условие за поддържане на живота на функциониращия организъм. В тази глава ще разгледаме основните механизми на такъв обмен. В следващите глави тези механизми ще бъдат разгледани във връзка с нервната клетка и нейните функции;
същите механизми обаче са в основата на функционирането на всички други органи.
Дифузия.Най-простият процес на преместване на вещество е дифузията. В разтвори (или газове) атомите и молекулите се движат свободно и разликата в концентрациите се балансира чрез дифузия. Да разгледаме два обема, пълни с течност или газ (фиг. 1.3), в които веществата имат концентрации c1 и c2 и разделени от слой с повърхност A и дебелинад. Потокът на материята m във време t описано Първият закон за дифузия на Фик:
дм/ dt= дА/ д ( ° С 1 –С 2)=дА/ дд ° С(1)
където D е коефициентът на дифузия, който е постоянен за дадено вещество, разтворител и температура. В по-общ вид, за разликата в концентрацията dc на разстояние dx
dm/dt= -D A dc/dx,(2)
потокът през секция А е пропорционален на градиента на концентрация dc/dx . Знакът минус се появява в уравнението, тъй като промяната в концентрацията в посока x е отрицателна.
Дифузията е най-важният процес, чрез който повечето от молекулите във водните разтвори се движат на къси разстояния. Това важи и за тяхното движение в клетката, доколкото дифузията не е възпрепятствана от мембрани. Много вещества могат свободно да дифундират през липидните мембрани, особено вода и разтворени газове като O 2 и CO 2 . мастноразтворим

Ориз. 1.3.Количествена схема на дифузия. Двете пространства са разделени от слой с дебелинади площ НО. C; - висока концентрация на частици в лявата част на обема, C:, - ниска концентрация на частици в дясната части, розова повърхносте концентрационният градиент в дифузионния слой. Дифузионен поток dm/dt – вж уравнение (1)
веществата също дифундират добре през мембраните; това важи и за относително малки полярни молекули, като етанол и урея, докато захарите преминават през липидния слой трудно. В същото време липидните слоеве са практически непроницаеми за заредени молекули, включително дори за неорганични йони. За неелектролити, дифузионното уравнение (1) обикновено се трансформира чрез комбиниране на характеристиките на мембраната и дифузиращото вещество в едно параметър-пропускливост (P):
dm/dt=P Aд ° С.(3)
На фиг. 1.4 в сравнение пропускливост (P) на липидната мембрана за различни молекули.
Дифузия през мембранни пори . Плазмената мембрана (и други клетъчни мембрани) са пропускливи не само за вещества, дифундиращи през липидния слой, но също и за много йони, захари, аминокиселини и нуклеотиди. Тези вещества преминават през мембраната през порите, образувани от транспортни протеини,вградени в мембраната. Вътре в такива протеини има пълен с вода канал с диаметър по-малък от 1 nm, през който малки молекули могат да дифундират. Те се движат по градиент на концентрация и ако носят заряд, тогава движението им през каналите също се регулира от мембранния потенциал. Мембранните канали са относително селективни

Ориз. 1.4.Пропускливост на изкуствени липидни двойни слоеве за различни вещества
по отношение на вида на молекулите, които могат да преминават през тях, има например калиеви, натриеви и калциеви канали, всеки от които е непроницаем за почти всеки йон, освен определен. Такава селективностпоради заряда или структурата на местата на свързване в стените на канала, което улеснява транспортирането на специфична молекула и предотвратява проникването на други вещества през канала (фиг. 1,5, А) .
Зад поведението мембранни йонни каналиЛесно е да се наблюдава, тъй като токът, възникващ по време на движението на йони, може да бъде измерен и дори за един канал. Показано е, че каналите спонтанно и с висока честота променят състоянието си от отворено към затворено. Калиевият канал се характеризира с токови импулси с амплитуда около 2 pA (2 10 -12 A) и продължителност няколко милисекунди (виж фиг. 2.12, стр. 37) [3]. През този период през него преминават десетки хиляди йони. Преходът на протеините от една конформация към друга се изследва чрез рентгенова дифракция, Мьосбауерова спектроскопия и ядрено-магнитен резонанс (ЯМР). Следователно протеините са силно динамични, подвижни структури и каналът през протеин не е просто твърда, пълна с вода тръба (Фигура 1.5A), а лабиринт от бързо движещи се молекулярни групи и заряди. Тази динамична реакция на канала се отразява в енергиен профил на канала,показано на фиг. 1.5, B. Тук абсцисата показва дължината на канала от външен разтвор с йонна концентрация C 0 и потенциал 0 до вътрешен разтвор с концентрация C 1 и потенциал E. Оста y

Ориз. 1.5.А. Схема на протеин, който образува калиев канал, вграден в липидния двуслой на плазмената мембрана. Четири отрицателни заряда са фиксирани на "стената" на канала. Б. Схематичен енергиен профил на канала, показан на фиг. A. По оста y се нанасят стойностите на кинетичната енергия, необходима за преминаване на канала; по оста на абсцисата, разстоянията между вътрешната и външната повърхност на мембраната. Енергийните минимуми съответстват на местата на свързване на положително заредени йони с фиксирани отрицателни заряди в стената на канала. Енергийните максимуми съответстват на дифузионни препятствия в канала. Предполага се, че конформацията на белтъка на канала спонтанно осцилира; опциите за енергийни профили са показани с плътни и пунктирани линии; тези трептения значително улесняват свързването на йони при преодоляване на енергийната бариера (но с промени)
показани са енергийните нива на йона в местата на свързване на канала; пикът в графиката представлява бариерата на пропускливостта, която йонната енергия трябва да преодолее, за да проникне в канала, а „понижаването“ на графиката представлява относително стабилно състояние (свързване). Въпреки препятствието на енергийния пик, йонът може да проникне в канала, ако енергийният профил се движи спонтанно; по този начин йонът може изведнъж да се появи "от другата страна" на енергийния пик и да продължи да се движи в клетката. В зависимост от заряда, размера и степента на хидратация на йона и способността му да се свързва със структурите на стените на канала, енергийният профил на канала варира за различните йони, което може да обясни селективността на отделните видове канали.
Дифузионно равновесие на йони . Дифузията на различни йони през мембранните канали трябва да доведе до елиминиране на разликите в концентрациите между екстра- и вътреклетъчната среда. Въпреки това, както се вижда от табл. 1.1, такива разлики продължават, така че трябва да има такива равновесиемежду дифузията и други транспортни процеси през мембраната. Следващите два раздела се занимават с начините, по които се установява такова равновесие. В случай на йони дифузионното равновесие се влияе от техния заряд. Дифузията на незаредени молекули се осигурява от разликата в концентрацията DC , а когато концентрациите се изравнят, действителният транспорт спира. Заредените частици са допълнително засегнати от електрическото поле. Например, когато калиев йон излезе от клетката по нейния концентрационен градиент, той носи един положителен заряд. По този начин вътреклетъчната среда става по-отрицателно заредена, което води до потенциална разлика през мембраната. Вътреклетъчният отрицателен заряд не позволява на нови калиеви йони да напуснат клетката, а тези йони, които въпреки това напускат клетката, допълнително ще увеличат заряда на мембраната. Потокът от калиеви йони спира, когато действието на електрическото поле компенсира дифузионното налягане поради разликата в концентрациите. Йоните продължават да преминават през мембраната, но в равни количества в двете посоки. Следователно за дадена разлика в концентрациите на йони върху мембраната съществува равновесен потенциалЕйон при което преминаването на йони през мембраната спира. Равновесният потенциал може лесно да се определи с помощта на уравнения на Нернст:
Ейон= Рт/ zФ* вътрешенВън/ C в(4)
където Р е газовата константа, T е абсолютната температура, z е йонната валентност (отрицателна за аниони)Вън е концентрацията на извънклетъчни йони, C в е вътреклетъчната концентрация на йона,Ф Числото на Фарадей. Ако заместим константи в уравнението, тогава при телесна температура (T = 310 K) равновесният потенциал за калиеви йони E K е равно на:
Ек= –61 мB дневник /(5)
Ако [ K + out ]/[ K + in ] = 39, както следва от табл. 1.1, тогава
Ek= -61 m B log 39= -97 mV.
Всъщност беше установено, че всички клетки имат мембранен потенциал;в мускулните клетки на бозайници нивото му е около -90 mV. В зависимост от условията и относителните концентрации на йони, клетките могат да имат мембранен потенциал в диапазона от -40 до -120mV. За клетката в примера по-горе (Таблица 1.1) потенциал за почивка,равно на приблизително -90 mV, показва, че потоците на калиеви йони през мембранните канали са приблизително в равновесие. Това не е изненадващо, тъй като отвореното състояние на калиеви канали в покойната мембрана е най-вероятно; мембраната е най-пропусклива за калиеви йони. Мембранният потенциал обаче се определя и от потоците на други йони.
Лекотата, с която незаредените частици могат да дифундират през мембраната, се определя количествено в уравнение (3). Пропускливост за заредени частици описано с малко по-сложно уравнение:
П= м Рт/ dF(6)
където ме подвижността на йона в мембраната,д - дебелина на мембраната, a R, T и F известни термодинамични константи. Стойностите на пропускливостта за различни йони, определени по този начин, могат да се използват за изчисляване на мембранния потенциалЕм когато калиеви, натриеви и хлоридни йони преминават през мембраната едновременно (с пропускливост P K, P Na и P Cl съответно). Предполага се, че потенциалът пада равномерно в мембраната, така че силата на полето е постоянна. В този случай се прилага Уравнение на Голдман или уравнение на постоянно поле :
Еm= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
За повечето клетъчни мембрани PК около 30 пъти по-висока от Rна (вижте също раздел 1.3). Относителна стойност PCl варира значително; за много мембрани PCl малък в сравнение с RК , но за други (например в скелетните мускули) PCl , много по-високо от RК.
Активен транспорт, натриева помпа . Предишният раздел описва пасивната дифузия на йони и получения мембранен потенциал при дадени вътреклетъчни и извънклетъчни йони. В резултат на този процес обаче концентрацията на йони вътре в клетката не се стабилизира автоматично, тъй като мембраната
потенциалът е малко по-електроотрицателен от E K , и много повече в сравнение с Ена (около +60 mV). Поради дифузията, вътреклетъчните концентрации на йони, най-малко калий и натрий, трябва да се изравнят с извънклетъчните. Стабилността на йонния градиент се постига чрез активен транспорт: мембранните протеини транспортират йони през мембраната срещу електрически и (или) концентрационни градиенти, като за това изразходват метаболитна енергия. Най-важният процес на активен транспорт е работата Na/K - помпа, която съществува в почти всички клетки;
помпата изпомпва натриеви йони от клетката, като същевременно изпомпва калиеви йони в клетката. Това осигурява ниска вътреклетъчна концентрация на натриеви йони и високо съдържание на калий (Таблица 1.1). Концентрационният градиент на натриевите йони върху мембраната има специфични функции, свързани с предаването на информация под формата на електрически импулси (виж раздел 2.2), както и с поддържането на други активни транспортни механизми и регулиране на клетъчния обем (виж по-долу). Ето защо не е изненадващо, че повече от 1/3 от енергията, консумирана от клетката, се изразходва за Na / K помпата, а в някои от най-активните клетки до 70% от енергията се изразходва за нейната работа.
Транспортният протеин Na/K е АТФаза. На вътрешната повърхност на мембраната той разгражда АТФ до АДФ и фосфат (фиг. 1.6). Енергията на една молекула АТФ се използва за транспортиране на три натриеви йона от клетката и едновременно два калиеви йона в клетката, т.е. общо един положителен заряд се отстранява от клетката за един цикъл. Така Na/K помпата е електрогенен(създава електрически ток през мембраната), което води до увеличаване на електроотрицателността на мембранния потенциал с приблизително 10 mV. Транспортният протеин извършва тази операция с висока скорост: от 150 до 600 натриеви йони в секунда. Аминокиселинната последователност на транспортния протеин е известна, но механизмът на този сложен обменен транспорт все още не е ясен. Този процес е описан с помощта на енергийните профили на преноса на натриеви или калиеви йони от протеини (фиг. 1.5.5). По естеството на промяната в тези профили, свързана с постоянни промени в конформацията на транспортния протеин (процес, който изисква енергия), може да се съди за стехиометрията на обмена: два калиеви йона се обменят с три натриеви йона.
Na/K-помпа, като изолирана Na+ /K + -зависима мембранна АТФаза, специално инхибирана от сърдечния гликозид уабаин (строфантин). Тъй като работата на Na/K помпата е многоетапна химическа реакция, тя, както всички химични реакции, до голяма степен зависи от температурата, което

Ориз. 1.6.Схема на Na/K-помпа-АТФаза (потопена в липидния бислой на плазмената мембрана), която в един цикъл извежда три Na + йона от клетката срещу градиенти на потенциал и концентрация и внася два K йона в клетката + . По време на този процес една молекула АТФ се разделя на ADP и фосфат. На диаграмата АТФазата е показана като димер, състоящ се от голяма (функционална) и малка субединица; в мембраната съществува като тетрамер, образуван от две големи и две малки субединици
показано на фиг. 1.7. Тук потокът на натриеви йони от мускулните клетки е показан във времето; това е практически еквивалентно на потока от натриеви йони, медииран от работата на помпата Na/K, тъй като пасивният поток на натриеви йони спрямо концентрационните и потенциалните градиенти е изключително малък. Ако препаратът се охлади с около 18°C, тогава потокът от натриеви йони от клетката бързо ще намалее 15 пъти и веднага след нагряване ще се възстанови до първоначалното си ниво. Такова намаляване на потока на натриеви йони от клетката е няколко пъти по-голямо от това, което би съответствало на температурната зависимост на процеса на дифузия или проста химическа реакция. Подобен ефект се наблюдава, когато метаболитната енергия е изчерпана в резултат на отравяне с динитрофенол (DNP) (фиг. 1.7.5). Следователно потокът от натриеви йони от клетката се осигурява от енергийно зависима реакция - активна помпа. Друга характеристика на помпата, наред със значителна температурна и енергийна зависимост, е наличието на ниво на насищане (както при всички други химични реакции); това означава, че скоростта на помпата не може да се увеличава безкрайно с увеличаване на концентрацията на транспортираните йони (фиг. 1.8). Обратно, потокът на пасивно дифузиращо вещество нараства пропорционално на разликата в концентрациите в съответствие със закона за дифузия (уравнения 1 и 2).

Ориз. 1.7. А, БС активен транспорт + . Y-ос:поток от радиоактивен 24 Na + от клетката (имп./мин.). абсцисната ос:време от началото на експеримента. НО.Клетката се охлажда от 18,3°С до 0,5°С; поток Na+ излизането от клетката през този период се инхибира. Б.Потискане на потока на Na + от клетката с динитрофенол (DNF) при концентрация 0,2 mmol/l (както е изменено)
В допълнение към помпата Na / K, плазмената мембрана съдържа поне още една помпа - калций;тази помпа изпомпва калциеви йони (Ca 2+) от клетката и участва в поддържането на тяхната вътреклетъчна концентрация на изключително ниско ниво (Таблица 1.1). Калциевата помпа присъства с много висока плътност в саркоплазмения ретикулум на мускулните клетки, които натрупват калциеви йони в резултат на разграждането на АТФ молекулите (вижте Глава 4).
Ефекти на Na/K помпата върху мембранния потенциал и клетъчния обем . На фиг. 1.9 показва различните компоненти на мембранния ток и показва вътреклетъчните концентрации на йони, които

Ориз. 1.8.Съотношението между скоростта на транспортиране на молекули и тяхната концентрация (в точката на влизане в канала или в точката на свързване на помпата) по време на дифузия през канала или по време на помпения транспорт. Последният се насища при високи концентрации (максимална скорост, Vmax ) стойност по абсцисата, съответстваща на половината от максималната скорост на помпата ( Vmax /2), е равновесната концентрация ДА СЕ м

Ориз. 1.9.Диаграма, показваща концентрациите на Na+ , K+ и Cl- вътре и извън клетката и пътищата за проникване на тези йони в клетъчната мембрана (чрез специфични йонни канали или с помощта на Na/K помпа. При дадени концентрационни градиенти, равновесните потенциали E Na, E K и E C l - са равни на посочените, мембранния потенциалЕм = – 90 mV
гарантират тяхното съществуване. През калиеви канали се наблюдава изходящ поток от калиеви йони, тъй като мембранният потенциал е малко по-електроположителен от равновесния потенциал за калиеви йони. Общата проводимост на натриевите канали е много по-ниска от тази на калиеви канали; натриевите канали са отворени много по-рядко от калиевите канали в състояние на покой; обаче приблизително същият брой натриеви йони влизат в клетката, както калиеви йони я напускат, тъй като са необходими голяма концентрация и потенциални градиенти за дифузията на натриеви йони в клетката. Na/K помпата осигурява идеална компенсация за пасивни дифузионни токове, тъй като транспортира натриеви йони от клетката и калиеви йони в нея. По този начин помпата е електрогенна поради разликата в броя на зарядите, прехвърлени в и извън клетката, която при нормална скорост на своята работа създава мембранен потенциал от около 10 mV е по-електроотрицателно, отколкото ако се генерира само от пасивни йонни токове (виж уравнение 7). В резултат на това мембранният потенциал се доближава до потенциала на калиевото равновесие, което намалява изтичането на калиеви йони. Na активност/K-помпата се регулира от вътреклетъчната концентрация на натриеви йони. Скоростта на помпата се забавя, тъй като концентрацията на натриеви йони, които трябва да бъдат отстранени от клетката, намалява (фиг. 1.8), така че работата на помпата и потокът от натриеви йони в клетката се балансират взаимно, поддържайки вътреклетъчната концентрация на натриеви йони на ниво около 10 mmol / l.
За да се поддържа баланс между изпомпване и пасивни мембранни токове, са необходими много повече Na/K-помпани молекули, отколкото има канални протеини за калиеви и натриеви йони. Когато каналът е отворен, десетки хиляди йони преминават през него за няколко милисекунди (вижте по-горе) и тъй като каналът обикновено се отваря няколко пъти в секунда, през него преминават общо повече от 10 5 йона за това време. Един протеин на помпа движи няколкостотин натриеви йони в секунда, така че плазмената мембрана трябва да съдържа около 1000 пъти повече помпени молекули, отколкото молекулите на канала. Измерванията на каналните токове в покой показват средно един калиев и един натриев отворен канал на 1 µm 2 мембрана; от това следва, че в едно и също пространство трябва да присъстват около 1000 Na/K помпени молекули, т.е. разстоянието между тях е средно 34 nm; диаметърът на изпомпващия протеин като канален протеин е 8–10 nm. Така мембраната е достатъчно плътно наситена с изпомпващи молекули
Фактът, че потокът от натриеви йони в клетката и калиеви йони извън клетката се компенсира от работата на помпата, има друго следствие, което се състои в поддържане на стабилно осмотично налягане и постоянен обем.Вътре в клетката има висока концентрация на големи аниони, главно протеини (А - в Таблица 1.1), които не са в състояние да проникнат през мембраната (или да проникват много бавно) и поради това са фиксиран компонент вътре в клетката. За да се балансира зарядът на тези аниони, е необходим равен брой катиони. Поради действието на Na/K помпата тези катиони са основно калиеви йони. Значително повишаване на вътреклетъчната концентрация на йони може да настъпи само при повишаване на концентрацията на аниони поради потока на C1 - по градиента на концентрацията в клетката (Таблица 1.1), но мембранният потенциал противодейства на това. Входящ ток Cl- наблюдава се само до достигане на равновесния потенциал за хлоридните йони; това се наблюдава, когато градиентът на хлоридните йони е почти противоположен на градиента на калиеви йони, тъй като хлоридните йони са отрицателно заредени (уравнение 4). По този начин се установява ниска вътреклетъчна концентрация на хлоридни йони, съответстваща на ниска извънклетъчна концентрация на калиеви йони. Резултатът е ограничаване на общия брой йони в клетката. Ако мембранният потенциал спадне, когато Na/K помпата е блокирана, например по време на аноксия, тогава равновесният потенциал за хлоридните йони намалява и съответно се увеличава вътреклетъчната концентрация на хлоридните йони. Възстановявайки баланса на зарядите, калиевите йони също влизат в клетката; общата концентрация на йони в клетката се увеличава, което повишава осмотичното налягане; това принуждава водата да влезе в клетката. Клетката набъбва. Вижда се това подуване in vivo в условия на липса на енергия.
градиент на концентрация на + като движеща сила за мембранния транспорт . Значението на Na/K-помпата за клетката не се ограничава до стабилизирането на нормалните K+ и Na+ градиенти през мембраната. Енергията, съхранявана в мембранния градиент Na+ , често се използва за осигуряване на мембранен транспорт за други вещества. Например, на фиг. 1.10 показва "symport" Na+ и захарните молекули в клетката. Мембранният транспортен протеин пренася захарната молекула в клетката дори срещу градиента на концентрация, докато Na + се движи по градиента на концентрация и потенциал, осигурявайки енергия за транспорт на захари.Такъв транспорт на захари зависи изцяло от наличието на висок градиент Na+ ; ако вътреклетъчната концентрация Na+ нараства значително, тогава пренасянето на захарите спира. За различни° С Ахаров, има различни симпортни системи. Транспорт на аминокиселинив клетка, подобна на транспортната° С Ахаров, показан на фиг. 1,10; той също се осигурява от градиента Na+ , има поне пет различни системи за симпорт, всяка от които е специализирана за една група свързани аминокиселини.
В допълнение към системите за символи, има също "антипорт".Един от тях, например, прехвърля един калциев йон извън клетката в един цикъл в замяна на три входящи натриеви йона (фиг. 1.10). Енергия за транспорт Ca 2+се образува поради навлизането на три натриеви йона по градиента на концентрация и потенциал. Тази енергия е достатъчна (при потенциал на покой) за поддържане на висок градиент на калциевите йони (от по-малко от 10–7 mol/l вътре в клетката до около 2 mmol/l извън клетката).
Ендо- и екзоцитоза . За определени вещества, които влизат в клетката или трябва да бъдат отстранени

Ориз. 1.10.Протеините, вградени в липидния бислой на мембраната, медиират вноса на глюкоза и Na+ в клетката, както и Ca2+/Na+ -антипорт, при който движещата сила е градиентът на Na + върху клетъчната мембрана
от него няма транспортни канали; такива вещества включват, например, протеини и холестерол. Те могат да преминат през плазмената мембрана в везикулиили мехурчета,чрез ендо- и екзоцитоза. На фиг. 1.11 показва основните механизми на тези процеси. По време на екзоцитозата определени органели (вижте по-долу) образуват везикули, пълни с вещество, което трябва да бъде отстранено от клетката, като хормони или извънклетъчни ензими. Когато такива везикули достигнат до плазмената мембрана, тяхната липидна мембрана се слива с нея, като по този начин позволява на съдържанието да избяга във външната среда. При обратния процес, ендоцитоза, плазмената мембрана инвагинира, образувайки ямка, която след това се задълбочава и затваря, образувайки вътреклетъчен везикул, пълен с извънклетъчна течност и някои макромолекули. За да се осигури това мембранно сливане и затваряне на везикула, контрактилните елементи на цитоскелета действат във връзка със самите мембрани (виж по-долу). Ендоцитозата не винаги включва просто улавяне на извънклетъчната среда в клетката. Клетъчната мембрана съдържа, често организирани в специализирани групи, специфични рецептори за макромолекули като инсулин или антигени. След като тези макромолекули се свържат с техните рецептори, настъпва ендоцитоза в областта на мембраната около рецептора и макромолекулата се транспортира селективно в клетката (фиг. 1.12, B).
Ендо- и екзоцитозата се появяват непрекъснато в клетките. Количеството циркулиращ мембранен материал е значително; в рамките на 1 час макрофагът абсорбира под формата на везикули, два пъти по-голяма от повърхността на цитоплазмената му мембрана. В повечето клетки оборотът на мембранния материал не е толкова интензивен, но все пак трябва да бъде значителен.

Ориз. 1.11.екзоцитоза и ендоцитоза. нагоре:вътреклетъчният везикул се слива с липидния бислой на плазмената мембрана и се отваря в извънклетъчното пространство. Този процес се нарича екзоцитоза. На дъното:плазмената мембрана инвагинира в малка област и отделя везикула, пълна с извънклетъчен материал. Този процес се нарича ендоцитоза.
1.3. Транспорт на вещества в клетката
Ендо- и екзоцитозата са не само процесите на транспортиране на вещества през клетъчната мембрана, но и процесите на мембранен обмен – структурните компоненти на самата клетка. Предмет на разглеждане в този раздел са други подобни транспортни процеси в клетката и нейните органели.

Ориз. 1.12. A-BСхема на процеси, включително екзо- и ендоцитоза. НО.Протеинът, синтезиран в гранулирания ендоплазмен ретикулум, се транспортира чрез апарата на Голджи до плазмената мембрана, където се секретира чрез екзоцитоза. Б.Холестеролът, свързан с LDL (липопротеини с ниска плътност) частици, се прикрепя към плазмената мембрана, индуцира образуването на ендоцитна везикула в този участък на мембраната и се транспортира до лизозомите, където се освобождава. INИзвънклетъчен материал, уловен по време на ендоцитоза (на фигурата на дясно),транспортиран през клетката във везикули или везикули и освободен чрез екзоцитоза (на фигурата наляво)
Дифузия . Естествено, в цитозола разликата в концентрацията се елиминира чрез дифузия; същото важи и за течностите, съдържащи се в органелите. Поради високата концентрация на разтворен протеин, дифузията тук е много по-бавна, отколкото във вода. Липидните мембрани - около клетката и в органелите - са двуизмерни течности, в които се осъществява дифузия. Липидите в мембранния двуслой дифундират в собствения си слой, рядко преминавайки от един в друг. Потопените в тях протеини също са доста подвижни; те се въртят около ос, перпендикулярна на мембраната, или дифундират странично с много различни дифузионни константи, 2–10 000 пъти по-бавни от фосфолипидите. Така че, ако някои протеини се движат свободно в липидния слой и със същата скорост като самите липидни молекули, тогава други са закотвени, т.е. доста силно свързан с цитоскелета. В мембраната има "постоянни" агрегати от специфични протеини, като пре- и постсинаптични структури на нервните клетки. Свободно движещите се протеини могат да бъдат демонстрирани чрез свързването им с флуоресцентни багрила, които се индуцират да светят чрез кратко осветяване на малка площ от мембраната с къси светкавици. Такива експерименти показват, че за по-малко от 1 минута протеините, свързани с багрилото, се разпределят равномерно върху мембраната на разстояния до 10 μm.
Активен транспорт в мембраните на органелите .
Процесите на активен транспорт, които играят жизненоважна роля за функционирането на плазмената мембрана, също протичат вътре в клетката, в мембраните на органелите. Специфичното съдържание на различни органели се създава отчасти чрез вътрешен синтез и отчасти чрез активен транспорт от цитозола. Един пример за последното е Ca 2+ помпата, спомената по-горе в саркоплазмения ретикулум на мускулните клетки. Особено интересно е, че в случай на синтез на АТФ в митохондриите, противоположният принцип важи за това, което се случва в АТФазните помпи на плазмената мембрана (фиг. 1.6). По време на синтеза на АТФ окислителният метаболизъм води до образуването на стръмен градиент H+ върху вътрешните мембрани. Този градиент е движещата сила за процес, който е обратен на цикъла на изпомпване на активен молекулен транспорт: H + йони се движат през мембраната по градиент, а енергията, освободена в резултат на това, осигурява синтеза на ATP от ADP и фосфат . Полученият АТФ от своя страна осигурява енергия на клетката, включително за активен транспорт.
Транспортиране във везикули . Клетката има голям брой органели и свързани везикули (фиг. 1.1). Тези органели, и особено везикули, са в постоянно движение, пренасяйки съдържанието си до други органели или до плазмената мембрана. Везикулите могат също да мигрират от клетъчната мембрана към органелите, както при ендоцитозата.
процес протеинова секрецияпоказано на фиг. 1.12 НО.Протеинът се синтезира близо до клетъчното ядро върху рибозоми, свързани с ендоплазмения ретикулум (т.нар. гранулиран, или груб, ендоплазмен ретикулум); Веднъж попаднал в ендоплазмения ретикулум, протеинът се пакетира в транспортни везикули, които се отделят от органелата и мигрират към апарата на Голджи. Тук те се сливат с резервоарите на апарата на Голджи, където протеинът се модифицира (т.е. превръща се в гликопротеин). В краищата на цистерните везикулите отново се отделят. Секреторните везикули, носещи модифицирания протеин, се придвижват към плазмената мембрана и освобождават съдържанието си чрез екзоцитоза.
Друг пример за транспортен път в клетка е показан на фиг. 1,12, В; е усвояването на холестерола от клетката. Холестеролът, транспортиран в кръвта, се свързва главно с протеини, като частици "липопротеини с ниска плътност"(LNP). Тези частици се прикрепят към специфични LDL рецепторни места на мембраната, където възниква ендоцитоза и LDL се транспортира в клетката в "покрити" везикули. Тези везикули се сливат, за да образуват ендозоми и губят своя „звън“ по време на този процес. Ендозомите от своя страна се сливат с първични лизозоми, съдържащи предимно хидролитични ензими, за да образуват вторични, по-големи лизозоми. При тях холестеролът се освобождава от LDL частици и дифундира в цитозола, където става достъпен, например, за синтеза на липидни мембрани. Везикулите, които не съдържат LDL, също се отделят от ендозомите, които се придвижват по специален начин към плазмената мембрана и се сливат с нея, връщайки мембранния материал и вероятно LDL рецепторите. От момента, в който LDL частицата се свърже с мембраната, минават 10-15 минути, докато холестеролът се освободи от вторичната лизозома. Нарушенията в свързването и усвояването на LDL, т.е. в доставянето на холестерол в клетката, играят решаваща роля в развитието на сериозно и широко разпространено заболяване, атеросклероза („втвърдяване“ на артериите).
Има много други транспортни маршрути, подобни на показаните на фиг. 1.11 и 1.12, А, с помощта на които се движат специфични везикули в клетката. Не е известно как точно се движат, но елементи от цитоскелета вероятно участват в този процес. Везикулите могат да се плъзгат по микротубулите, като в този случай енергията за движение изглежда се осигурява от свързания с везикулите протеин, ATPase (виж по-долу). Остава напълно неразбираемо колко различни везикули, движещи се един след друг във всички посоки, стигат до местоназначението си. Те очевидно трябва да бъдат „маркирани“ по такъв начин, че да бъдат разпознати от транспортната система и превърнати в целенасочено движение.
Транспортиране чрез образуване и разрушаване на органели . Досега ние разглеждахме ендо- и екзоцитозата като процеси на транспортиране на съдържанието на везикулите. Има и друг аспект на тези процеси, който се състои във факта, че насоченото отстраняване на плазмената мембрана в една област от клетъчната повърхност чрез ендоцитоза и, напротив, добавянето й към друга чрез екзоцитоза, премества значителни участъци от мембрана (фиг. 1.12.E), което дава възможност на клетката, например, да образува израстък или да се премести.
Подобни пренареждания са характерни и за цитоскелета, особено за микрофиламентите и микротубулите (фиг. 1.1). Микрофиламентисе състоят предимно от F-актин протеинкойто е способен да се сглобява във влакнести снопчета в резултат на полимеризация на мономера от цитозола. Снопчетата са поляризирани, тоест често растат само от единия край, натрупвайки нови актинови молекули, докато другият край е инертен или тук става разглобяване. Поради този поляризиран растеж, микрофиламентите се движат ефективно и структурата на тяхната мрежа може да се промени. Преходът на актина от деполимеризирано състояние (sol) в организирано състояние (гел) може да се случи много бързо под въздействието на други протеини или промени в концентрацията на йони (вижте по-долу). Има и протеини, които карат актиновите нишки да се разпадат на къси фрагменти. Тънките израстъци на много клетки - филоподии - съдържат централен сноп актин (фиг. 1.1), а различните движения на филоподиите вероятно се дължат на актинови преходи: полимеризация - деполимеризация.
микротубулисъщо често се подлагат на подобни движения. Механизмът на тези движения е подобен - полимеризацията на тубулин от цитозола по такъв начин, че единият от краищата на микротубулата расте, докато другият или не се променя, или се разглобява там. По този начин, микротубулата, чрез подходящо добавяне или отстраняване на материал, може да се движи през цитозола.
Активни движения на цитоскелета . Промени в цитоскелетните структури могат да възникнат в резултат както на активни движения, така и на пренареждания, описани по-горе. В много случаи движението на микротубулите и актиновите филаменти се задвижва от контрактилни протеини, които свързват филаментите или тубулите и могат да ги преместват един спрямо друг. катерици миозин и динеинприсъства в цитозола на всички клетки в относително високи концентрации; те са елементите, които превръщат енергията в движение в специализирани клетки (мускул) и органели (реснички). В мускулните клетки миозинът образува дебели нишки, ориентирани успоредно на актиновите нишки. Молекулата на миозина със своята „глава“ се прикрепя към актиновата нишка и използвайки енергията на АТФ, измества миозина по протежение на молекулата на актина.След това миозинът се отделя от актина. Наборът от много такива цикли свързване-изключване води до макроскопично свиване на мускулните влакна(Гл. 4). Dynein играе подобна роля в движението на микротубулите по време на операция на ресничките (фиг. 1.1). В цитоплазмата на неспециализирани клетки миозинът и динеинът не образуват редовни влакна, а в повечето случаи малки групи от молекули. Дори под формата на такива малки агрегати, те са в състояние да движат актинови филаменти или микротубули. Ориз. 1.13 илюстрира този процес, когато противоположно поляризирани миозинови молекули също са прикрепени към две актинови нишки, поляризирани в противоположни посоки. Главните групи на миозина се огъват към опашката на молекулата, консумирайки АТФ, а двете актинови нишки се изместват в обратна посока, след което миозинът се отделя от тях. Движенията от този вид, по време на които енергията на АТФ се превръща в механична работа, могат да променят формата на цитоскелета и следователно на клетките, както и да осигурят транспорт на органели, свързани с цитоскелета.
Процесите на вътреклетъчен транспорт могат да бъдат най-ясно демонстрирани върху аксона на нервната клетка. транспорт на аксонобсъдени тук подробно, за да илюстрират събития, които вероятно се случват по подобен начин в повечето клетки. Аксонът с диаметър само няколко микрона може да бъде дълъг един метър или повече и ще са необходими години, докато протеините се придвижат чрез дифузия от ядрото до дисталния край на аксона. Отдавна е известно, че когато някоя част от аксона претърпи свиване, проксималната част на аксона се разширява. Изглежда, че центробежният поток е блокиран в аксона. Такава поток-бърз транспорт на аксон можесе демонстрира чрез движението на радиоактивни маркери, както в експеримента, показан на фиг. 1.14. Радиоактивно белязан левцин се инжектира в ганглия на дорзалния корен и след това от 2-ия до 10-ия час се измерва радиоактивността в седалищния нерв на разстояние 166 mm от телата на невроните. В продължение на 10 часа пикът на радиоактивност на мястото на инжектиране не се промени значително. Но вълната на радиоактивност се разпространява по аксона с постоянна скорост от около 34 mm за 2 часа, или 410 mm/ден. Доказано е, че при всички неврони на хомойотермични животни бързият транспорт на аксон се осъществява с еднаква скорост и няма забележими разлики между тънките, немиелинизирани влакна и най-дебелите аксони, както и между двигателните и сетивните влакна. Видът на радиоактивния маркер също не влияе върху скоростта на бърз аксонален транспорт; Различни радиоактивни вещества могат да служат като маркери.

Ориз. 1.13.Немускулен миозинов комплекс с определена ориентация може да се свърже с актинови нишки с различна полярност и, използвайки енергията на АТФ, да ги измести един спрямо друг.
молекули, като различните аминокиселини, които са включени в телесните протеини на неврон. Ако анализираме периферната част на нерва, за да определим естеството на носителите на пренасяната тук радиоактивност, тогава такива носители се намират главно в протеиновата фракция, но също и в състава на медиаторите и свободните аминокиселини. Като знаем, че свойствата на тези вещества са различни и особено размерите на техните молекули са различни, можем да обясним постоянната скорост на транспорт само с общия за всички транспортен механизъм.
Описано по-горе бърз транспорт на аксоне антерограднанасочени далеч от тялото на клетката. Доказано е, че някои вещества се придвижват от периферията към клетъчното тяло с помощта на ретрограден транспорт.Например, ацетилхолинестеразата се транспортира в тази посока със скорост, два пъти по-ниска от скоростта на бързия аксонален транспорт. Маркер, често използван в невроанатомията - пероксидаза от хрян - също се движи ретроградно. Ретроградният транспорт вероятно играе важна роля в регулирането на протеиновия синтез в клетъчното тяло. Няколко дни след трансекцията на аксона се наблюдава хроматолиза в клетъчното тяло, което показва нарушение на протеиновия синтез. Времето, необходимо за хроматолиза, корелира с продължителността на ретрограден транспорт от мястото на трансекция на аксона до тялото на клетката. Подобен резултат също предлага обяснение за това нарушение - нарушава се предаването от периферията на „сигналната субстанция“, която регулира протеиновия синтез. Очевидно основните "превозни средства", използвани за бърз аксон
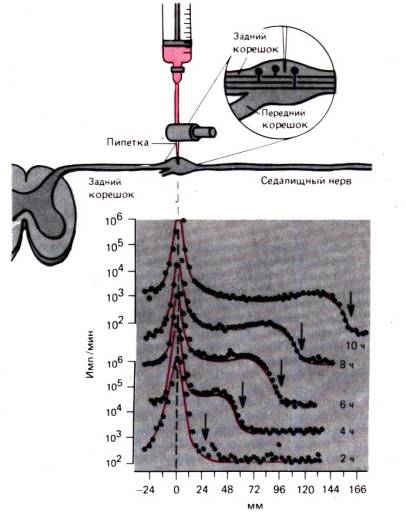
Ориз. 1.14.Експеримент, демонстриращ бърз транспорт на аксон в сензорните влакна на седалищния нерв на котка. Белязан с тритий левцин се инжектира в ганглия на дорзалния корен и радиоактивността в ганглия и сетивните влакна се измерва 2, 4, 6, 8 и 10 часа след инжектирането. (долната част на фигурата).от абсцисаразстоянието от ганглия до участъците на седалищния нерв, където се извършва измерването, се отлага. По оста y, само за горната и долната криви, радиоактивността (imp./min) е нанесена в логаритмична скала. "Вълна" на повишена радиоактивност (стрелки)движи се със скорост от 410 мм/ден (от )
транспорт са везикули (везикули) и органели,като митохондриите, съдържащи вещества, които трябва да бъдат транспортирани. Движението на най-големите везикули или митохондриите може да се наблюдава с помощта на микроскоп in vivo . Такива частици правят кратки, бързи движения в една от посоките, спират, често се движат малко назад или встрани, спират отново и след това правят рязко движение в основната посока. 410 mm/ден съответстват на средна антероградна скорост от приблизително 5 μm/s; следователно скоростта на всяко отделно движение трябва да бъде много по-висока и ако вземем предвид размера на органелите, филаментите и микротубулите, тогава тези движения са наистина много бързи. Бързият транспорт на аксон изисква значителна концентрация на АТФ. Отрови като колхицин, разрушаващ микротубулите, също блокират бързия аксонален транспорт. От това следва, че в транспортния процес, който разглеждаме, везикулите и органелите се движат по микротубулите и актиновите филаменти; това движение се осигурява от малки агрегати от молекули динеин и миозин, действащи, както е показано на фиг. 1.13, използвайки енергията на АТФ.
Бързият транспорт на аксон може също да бъде включен патологични процеси.Някои невротропни вируси (например вируси на херпес или полиомиелит) проникват в аксона в периферията и се придвижват чрез ретрограден транспорт до тялото на неврона, където се размножават и упражняват токсичния си ефект. Тетанусният токсин, протеин, произведен от бактерии, който навлиза в тялото чрез кожни лезии, се поема от нервните окончания и се транспортира до тялото на неврона, където причинява характерни мускулни спазми. Известни са случаи на токсични ефекти върху самия аксонен транспорт, например излагане на индустриален разтворител акриламид. Освен това се смята, че патогенезата на бери-бери бери-бери и алкохолна полиневропатия включва нарушение на бързия аксонен транспорт.
В допълнение към бърз транспорт на аксон в клетката, има и доста интензивен бавен транспорт на аксон.Тубулинът се движи по аксона със скорост от около 1 mm/ден, докато актинът се движи по-бързо, до 5 mm/ден. Други протеини също мигрират с тези компоненти на цитоскелета; например, ензимите изглежда са свързани с актин или тубулин. Скоростите на движение на тубулин и актин са приблизително в съответствие със скоростите на растеж, установени за механизма, описан по-рано, когато молекулите са включени в активния край на микротубула или микрофиламент. Следователно този механизъм може да лежи в основата на бавен аксонален транспорт. Скоростта на бавния транспорт на аксони също приблизително съответства на скоростта на растеж на аксона, което очевидно показва ограниченията, наложени от структурата на цитоскелета върху втория процес.
В заключение на този раздел трябва да се подчертае, че клетките в никакъв случай не са статични структури, както изглеждат например на електронно-микроскопски снимки. плазмената мембранаи особено органелите са в постоянно бързо движение и постоянно преструктуриране;това е единствената причина те да могат да функционират. Освен това, това не са прости камери, в които протичат химични реакции, а високоорганизирани конгломерати от мембрани и влакна,при който реакциите протичат в оптимално организирана последователност.
1.4. Регулиране на клетъчните функции
Поддържането на отделната клетка като функционална единица до голяма степен се регулира от ядрото; изучаването на такива регулаторни механизми е предмет на клетъчната биология и биохимия. В същото време клетките трябва да променят функциите си в съответствие с условията на околната среда и нуждите на други клетки на тялото, т.е. те служат като обекти на функционална регулация. По-долу ще разгледаме накратко как тези регулаторни влияния действат върху плазмената мембрана и как достигат до вътреклетъчните органели.
Регулаторни ефекти върху клетъчната мембрана
Мембранен потенциал . В много случаи регулирането на клетъчните функции се осъществява чрез промяна на мембранния потенциал. Локалните промени в потенциала са възможни, когато: 1) ток от съседна клетъчна област или генериран от друга клетка протича през мембраната; 2) извънклетъчната концентрация на йони се променя (често [K + ]навън ); 3) мембранните йонни канали се отварят. Промените в мембранния потенциал могат да повлияят на конформацията на мембранните протеини, причинявайки по-специално отваряне или затваряне на канали. Както е описано по-горе, функционирането на някои мембранни помпи зависи от мембранния потенциал. Нервните клетки са специализирани да възприемат промените в мембранния потенциал като информация, която трябва да се обработва и предава (вижте Глава 2).
извънклетъчни регулаторни вещества . Най-важният регулаторен механизъм, включващ извънклетъчните вещества, е тяхното взаимодействие със специфични рецептори на плазмената мембрана или вътре в клетката. Тези вещества включват синаптични медиатори, които предават информация между нервните клетки, локални агенти и вещества, които циркулират в кръвта и достигат до всички клетки на тялото, като хормони и антигени. синаптични невротрансмитериса малки молекули, освободени от нервните окончания в синапса;
когато достигнат до плазмената мембрана на съседна постсинаптична клетка, те задействат електрически сигнали или други регулаторни механизми. Този въпрос е разгледан подробно в гл. 3.
Местни химични агенти често се секретира от специализирани клетки. Те дифундират свободно в извънклетъчното пространство, но тяхното действие е ограничено до малка група клетки поради бързото унищожаване на тези вещества, спонтанно или чрез действието на ензими. Един пример за освобождаване на такива агенти е освобождаването хистаминмастоцити при нараняване или имунен отговор. Хистаминът причинява релаксация на клетките на гладката мускулатура на съдовете, повишава пропускливостта на съдовия ендотел и стимулира сетивните нервни окончания, които медиират усещането за сърбеж. Други локални химични агенти се секретират от много други клетки. Типичните местни агенти са простагландини,съставляващи група от приблизително 20 производни на мастни киселини. Те се освобождават непрекъснато от широко разпространени клетки, но действат само локално, тъй като бързо се разрушават от мембранните фосфолипази. Различните простагландини имат широк спектър на действие: те могат да предизвикат свиване на гладкомускулните клетки, да причинят агрегация на кръвните тромбоцити (тромбоцити) или да инхибират развитието на жълтото тяло в яйчниците.
Обслужват други местни агенти растежни фактори.Най-известният нервен растежен фактор (NGF) за симпатиковите неврони, който е необходим за растежа и оцеляването на тези неврони по време на развитието in vivo или в клетъчна култура. Очевидно прицелните клетки за този клас неврони секретират NGF и по този начин осигуряват правилната инервация. Когато формират органи, клетките често трябва да „намерят пътя“ към целевите клетки, които могат да бъдат разположени на значителни разстояния. Съответно трябва да има много специализирани растежни фактори като NGF.
Хормони и антигени пренасяни от кръвта до всички клетки. Антигените предизвикват имунен отговор от клетки, които носят специфични антитела. Антигените обаче по правило са чужди вещества, които не се образуват в реагиращия организъм (за повече подробности вижте глава 18). Някои хормони, като инсулин или тироксин, засягат голямо разнообразие от типове клетки, докато други, като половите хормони, засягат само определени видове клетки. Хормоните са или пептиди, чието действие се задейства от свързването им с рецептор на клетъчната мембрана, или стероиди и тироксин, които дифундират през липидната мембрана и се свързват с вътреклетъчните рецептори. Стероидните хормони се свързват с ядрения хроматин, което води до транскрипция на определени гени. Произведените в резултат на това протеини предизвикват промяна в клетъчните функции, което е специфичното действие на хормоните. Въпросите, свързани с освобождаването и действието на хормоните, са разгледани подробно в гл. 17.
Вътреклетъчна комуникация, включваща втори пратеници
Регулаторните функции, описани по-горе, включват ефекти върху клетъчната мембрана. Информацията, получена от клетъчната мембрана, често трябва да предизвика реакция в органелите и се пренася към тях от различни вещества, известни като втори пратеници (за разлика от първите, идващи в клетката от външни източници). Изучаването на вторите медиатори се развива бързо и няма гаранция, че настоящото ниво на разбиране на проблема ще бъде достатъчно пълно. Тук ще засегнем три добре проучени медиатора: Ca 2+, cAMP и инозитол трифосфат.
калций.Най-простият вътреклетъчен медиатор е Ca 2+ йонът. Свободната му концентрация в покойна клетка е много ниска и възлиза на 10 -8 -10 -7 mol/l. Той може да влезе в клетката през специфични мембранни канали, когато са отворени, например, когато мембранният потенциал се промени (вижте Глава 2). Полученото увеличение на Ca 2+ предизвиква важни реакции в клетката, като свиване на миофибрилите, което е в основата на мускулната контракция (вижте Глава 4), или освобождаването на съдържащи невротрансмитери везикули от нервните окончания (вижте Глава 3) . И двете реакции изискват концентрация на Ca 2+ от приблизително 10–5 mol/l. Ca 2+, който има регулаторен ефект, може да се освобождава и от вътреклетъчни депа, като ендоплазмения ретикулум. Освобождаването на Ca 2+ от депото изисква участието на други посредници (виж, например, фиг. 1.16).
Цикличен аденозин монофосфат, cAMP. Напоследък беше доказано, че цикличният аденозин монофосфат (cAMP), производно на основния енергиен източник в тялото, АТФ, е важен втори пратеник. Сложната верига от реакции, показана на фиг. 1.15, започва от рецептора Rs върху външната повърхност на плазмената мембрана, която може да служи като специфично свързващо място за различни медиатори и хормони. След свързване със специфична "стимулираща" молекула Rs променя своята конформация; тези промени засягат протеина Gs върху вътрешната повърхност на мембраната по такъв начин, че става възможно да се активира последната чрез вътреклетъчен гуанозин трифосфат (GTP). активиран протеин Gs , от своя страна, стимулира ензим на вътрешната повърхност на мембраната, аденилат циклаза (АС), който катализира образуването на сАМР от АТФ. Водоразтворим cAMP и е медиаторът, който предава ефекта

Ориз. 1.15.Верига от реакции, включващи вътреклетъчния медиатор cAMP (цикличен аденозин монофосфат). Възбуждащите или инхибиторните външни сигнали активират мембранните рецептори R s или Ri . Тези рецептори регулират процеса на свързванег -протеини с вътреклетъчен GTP (гуанозин трифосфат), като по този начин стимулират или инхибират вътреклетъчната аденилатциклаза (AC). Усилващият ензим AC превръща аденозин трифосфат (АТФ) в cAMP, който след това се разцепва до AMP с помощта на фосфодиестер. Свободният сАМР дифундира в клетката и активира аденилат киназата (А-киназа), освобождавайки нейната каталитична субединица С, която катализира фосфорилирането на вътреклетъчните протеини, т.е. формира крайния ефект на извънклетъчния стимул. Схемата също така показва фармакологични лекарства и токсини, които предизвикват (+) или инхибират (-) някои реакции (но с промени)
стимулиране на извънклетъчния рецептор Rs към вътрешните структури на клетката.
Успоредно със стимулиращата верига от реакции, включващи Rs възможно свързване на инхибиторни медиатори и хормони към съответния рецептор R и което отново чрез GTP-активиран протеинг , инхибира AC и по този начин производството на cAMP. Дифундирайки в клетката, сАМР реагира с аденилат киназа (А-киназа); Това освобождава субединицата C, която катализира протеиновото фосфорилиране.Това фосфорилиране превръща протеините в тяхната активна форма и сега те могат да упражняват своето специфично регулаторно действие (например да предизвикат разграждане на гликоген). Тази сложна регулаторна система е изключително ефективна, тъй като крайният резултат е фосфорилирането на много протеини, т.е. регулаторният сигнал преминава през веригата с голям фактор на усилване. Външни медиатори, които се свързват с рецепторите R s и R i специфични за всеки от тях са изключително разнообразни. Адреналин, забъркване с R s или R i участва в регулирането на метаболизма на липидите и гликогена, както и в засиленото свиване на сърдечния мускул и в други реакции (виж глава 19). тироид-стимулиращ хормон, активиране Rs , стимулира секрецията на хормона тироксин от щитовидната жлеза, а простагландин I инхибира агрегацията на тромбоцитите. Инхибиращи ефекти, включително адреналин, медиирани чрез R и изразяващ се в забавяне на липолизата. По този начин, cAMP системата е многофункционална вътреклетъчна регулаторна система,които могат да бъдат прецизно контролирани от извънклетъчни стимулиращи и инхибиторни сигнални агенти.
Инозитол фосфат "IF з ". Вътреклетъчната система на втория медиатор, инозитол фосфат, е открита едва наскоро (фиг. 1.16). В този случай няма инхибиторен път, но има прилика със cAMP системата, при която ефектът от стимулиране на R рецептора се прехвърля към GTP-активирания G-протеин върху вътрешната повърхност на мембраната. На следващия етап обичайният мембранен липиден фосфатидилинозитол (PI), който преди това е получил две допълнителни фосфатни групи, се превръща в PI-дифосфат (FIF 2), който се разцепва от активирана фосфодиестераза (PDE) в инозитол трифосфат(IFz) и липид диацилглицерол(DAG). Инозитол трифосфатът е водоразтворим втори пратеник, който дифундира в цитозола. Действа основно чрез освобождаване на Ca 2+ от ендоплазмения ретикулум. Ca 2+ от своя страна действа като посредник, както е описано по-горе; например активира Са2+-зависимата фосфокиназа, която фосфорилира ензимите. Липидната субединица на DAG (фиг. 1.16) също носи сигнал, дифундиращ в липидната фаза на плазмената мембрана към С-киназата, разположена на нейната вътрешна повърхност, която се активира с участието на фосфатидилсерин като кофактор. Тогава С-киназата задейства фосфорилирането на протеини, превръщайки ги в активна форма.
Вътреклетъчната система на втория междинен интерферон 3 може също да се контролира от различни външни медиатори и хормони, включително ацетилхолин, серотонин, вазопресин и тироид-стимулиращ хормон; подобно на cAMP системата, тя се характеризира с различни вътреклетъчни ефекти. Възможно е тази система също да се активира от светлина в зрителния рецептор на окото и да играе централна роля във фототрансдукцията (вж. Глава 11). За първи път в индивидуалното развитие на организма рецепторът на IGF системата се активира от сперматозоидите, в резултат на което IGF участва в регулаторните реакции, които съпътстват оплождането на яйцеклетката.
Системите cAMP и IFz-DAG са много ефективни биологични подобрители.Те са

Ориз. 1.16.Верига от реакции, включващи вътреклетъчен междинен интерферон (инозитол трифосфат). Както в cAMP системата, извънклетъчният сигнал се медиира чрез протеинг, който в този случай активира фосфодиестераза (PDE). Този ензим разгражда фосфатидилинозин дифосфат (FIF) 2 ) в плазмената мембрана преди IFз и диацилглицерол (DAG); АКОз дифундира в цитоплазмата. Тук той предизвиква освобождаването на Ca 2+ от ендоплазмения ретикулум; повишаване на концентрацията на Ca 2+ в цитоплазмата ([Ca 2+ ] i ) активира протеин киназа, която фосфорилира и следователно активира ензими. Друг продукт, DAG, остава в мембраната и активира протеин киназа С (кофактор-фосфатидилсерин, PS). Протеин киназа С също така фосфорилира ензими, които медиират специфични действия, свързани със стимулиране на външния рецептор.Р . Клонове на веригата от реакции, включващи IFз и DAG могат да се активират независимо от йономицин и форболов естер, съответно (модифициран)
превръща реакцията между медиатора и рецептора на външната мембрана във фосфорилиране на много вътреклетъчни протеини, които след това могат да повлияят на различни клетъчни функции. Един от съществените аспекти на проблема е, че доколкото знаем днес, съществуват само тези две тясно свързани регулаторни системи от този тип, които се използват от множество външни медиатори за регулиране на различни вътреклетъчни процеси. В същото време тези регулаторни системи, включително Ca 2+, взаимодействат тясно една с друга, което им позволява да извършват фина регулация на клетъчните функции.
Уроци и ръководства
1. Албъртс IN., Брей Д., Люис Дж., Раф М., Робъртс ДА СЕ., Уотсън Дж.Д.Молекулярна биология на клетката, Ню Йорк и Лондон, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(ред.). Биология. Берлин, Хайделберг, Ню Йорк, Springer, 1983 г.
3. хил IN. Йонни канали на възбудимите мембрани. Съндърланд, Масачузетс, Sinauer Assoc., 1984.
4. Хопе В., Ломан В. Марки Х., Зиглер Х.(ред.). Биофизик. Берлин, Хайделберг, Ню Йорк, Springer, 1984.
5. Юнгерман ДА СЕ., Малер Х.биохимия. Берлин, Хайделберг, Ню Йорк, Springer, 1980 г.
6. Кандел Е.Р., Шварц-Дж.Х.,(ред.). Принципи на неврологичните науки, Ню Йорк, Амстердам, Оксфорд, Elsevier, 1985.
7. Шиблер т. H., SchmidtW.Анатомичен дез Menschen. Берлин, Хайделберг, Ню Йорк, Токио, Springer, 1983 г.
Оригинални статии и рецензии
8. Berridge M.J.Молекулната основа на комуникацията в клетката, Sci. Amer 253 124 134 (1985).
9. Беридж М. Дж., Ървайн Р. Ф.Инозитол трифосфат, нов втори пратеник в клетъчната сигнална трансдукция. Nature, 312, 315 321 (1984).
10. Bretscher M.S.Молекулите на клетъчната мембрана, Sci. Amer., 253, 124–134 (1985).
11. Даут Дж.Живата клетка като машина за преобразуване на енергия. Минимален модел на миокарден метаболизъм, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Ходжкин А.Л., Кац IN. Ефектът на натриевите йони върху електрическата активност на гигантския аксон на калмара. J Physiol. (Лонд.), 108, 37–77 (1949).
13. Ходжкин А.Л., Кейнс Р.Д.Активен транспорт на катиони в гигантски аксони от сепияи лолиго, J Physiol. (Лонд.), 128, 28–42 (1955).
14. По-дълъг П.Йонни канали с конформационни подсъстояния, Биофиз. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M.Екзоплазмен транспорт в нормални и патологични системи. В: Физиология и патология на аксоните, S.G. Waxman,Изд. Ню Йорк, Raven Press, 1978 г.
Каним ви да се запознаете с материалите и.
: целулозна мембрана, мембрана, цитоплазма с органели, ядро, вакуоли с клетъчен сок.Наличието на пластиди е основната характеристика на растителната клетка.
Функции на клетъчната стена- определя формата на клетката, предпазва от факторите на околната среда.
плазмената мембрана- тънък филм, съставен от взаимодействащи липидни и протеинови молекули, ограничава вътрешното съдържание от външната среда, осигурява транспортиране на вода, минерални и органични вещества в клетката чрез осмоза и активен трансфер, а също така отстранява отпадъчните продукти.
Цитоплазма- вътрешната полутечна среда на клетката, в която се намират ядрото и органелите, осигурява връзки между тях, участва в основните процеси на живот.
Ендоплазмения ретикулум- мрежа от разклоняващи се канали в цитоплазмата. Участва в синтеза на протеини, липиди и въглехидрати, в транспорта на вещества. Рибозоми - тела, разположени върху EPS или в цитоплазмата, състоят се от РНК и протеин, участват в синтеза на протеини. EPS и рибозомите са единен апарат за синтез и транспорт на протеини.
митохондриите-органели, отделени от цитоплазмата с две мембрани. В тях се окисляват органични вещества и с участието на ензими се синтезират молекули на АТФ. Увеличение на повърхността на вътрешната мембрана, върху която са разположени ензими, поради кристи. АТФ е органично вещество, богато на енергия.
пластиди(хлоропласти, левкопласти, хромопласти), тяхното съдържание в клетката е основна характеристика на растителния организъм. Хлоропластите са пластиди, съдържащи зеления пигмент хлорофил, който абсорбира светлинната енергия и я използва за синтезиране на органични вещества от въглероден диоксид и вода. Разграничаване на хлоропластите от цитоплазмата от две мембрани, многобройни израстъци - грана върху вътрешната мембрана, в която са разположени хлорофилни молекули и ензими.
комплекс Голджи- система от кухини, ограничени от цитоплазмата с мембрана. Натрупването на протеини, мазнини и въглехидрати в тях. Осъществяване на синтеза на мазнини и въглехидрати върху мембрани.
лизозоми- тела, отделени от цитоплазмата с една мембрана. Съдържащите се в тях ензими ускоряват реакцията на разделяне на сложни молекули на прости: протеини до аминокиселини, сложни въглехидрати до прости, липиди до глицерол и мастни киселини, а също така унищожават мъртвите части на клетката, цели клетки.
Вакуоли- кухини в цитоплазмата, пълни с клетъчен сок, място за натрупване на резервни хранителни вещества, вредни вещества; те регулират съдържанието на вода в клетката.
Ядро- основната част на клетката, покрита отвън с двумембрана, пробита от пори ядрена обвивка. Веществата навлизат в сърцевината и се отстраняват от нея през порите. Хромозомите са носители на наследствена информация за характеристиките на даден организъм, основните структури на ядрото, всяка от които се състои от една молекула ДНК в комбинация с протеини. Ядрото е мястото на синтеза на ДНК, i-RNA, r-RNA.

Наличието на външна мембрана, цитоплазма с органели, ядро с хромозоми.
Външна или плазмена мембрана- ограничава съдържанието на клетката от околната среда (други клетки, междуклетъчно вещество), състои се от липидни и протеинови молекули, осигурява комуникация между клетките, транспорт на вещества в клетката (пиноцитоза, фагоцитоза) и извън клетката.
Цитоплазма- вътрешната полутечна среда на клетката, която осигурява комуникация между ядрото и разположените в нея органели. Основните процеси на жизнена дейност протичат в цитоплазмата.
Клетъчни органели:
1) ендоплазмен ретикулум (ER)- система от разклонени тубули, участващи в синтеза на протеини, липиди и въглехидрати, в транспорта на вещества в клетката;
2) рибозоми- телата, съдържащи rRNA, са разположени върху ER и в цитоплазмата и участват в протеиновия синтез. EPS и рибозомите са единен апарат за протеинов синтез и транспорт;
3) митохондрии- "електростанции" на клетката, ограничени от цитоплазмата с две мембрани. Вътрешният образува кристи (гънки), които увеличават повърхността му. Ензимите върху кристите ускоряват реакциите на окисление на органичните вещества и синтеза на енергийно богати АТФ молекули;
4) комплекс Голджи- група от кухини, ограничени с мембрана от цитоплазмата, пълни с протеини, мазнини и въглехидрати, които се използват или в жизнените процеси, или се отстраняват от клетката. Мембраните на комплекса осъществяват синтеза на мазнини и въглехидрати;
5) лизозоми- тела, пълни с ензими, ускоряват реакциите на разграждане на протеини до аминокиселини, липиди до глицерол и мастни киселини, полизахариди до монозахариди. В лизозомите се унищожават мъртвите части на клетката, цели клетки и клетки.
Клетъчни включвания- Натрупване на резервни хранителни вещества: протеини, мазнини и въглехидрати.
Ядро- най-важната част от клетката. Покрит е с двумембранна мембрана с пори, през които едни вещества проникват в ядрото, а други влизат в цитоплазмата. Хромозомите са основните структури на ядрото, носители на наследствена информация за характеристиките на организма. Предава се в процеса на делене на майчината клетка към дъщерните клетки, а със зародишните клетки - към дъщерните организми. Ядрото е мястото на синтеза на ДНК, иРНК, рРНК.
Задачата:
Обяснете защо органелите се наричат специализирани структури на клетката?
Отговор:органелите се наричат специализирани клетъчни структури, тъй като изпълняват строго определени функции, наследствената информация се съхранява в ядрото, АТФ се синтезира в митохондриите, фотосинтезата протича в хлоропластите и др.
Ако имате въпроси относно цитологията, можете да помолите за помощ от
Третият етап от еволюцията е появата на клетката.
Молекулите на протеини и нуклеинови киселини (ДНК и РНК) образуват биологична клетка, най-малката единица на живота. Биологичните клетки са „градивните елементи“ на всички живи организми и съдържат всички материални кодове на развитие.
Дълго време учените смятаха структурата на клетката за изключително проста. Съветският енциклопедичен речник тълкува понятието клетка по следния начин: „Клетката е елементарна жива система, основа на структурата и живота на всички животни и растения“. Трябва да се отбележи, че терминът "елементарен" в никакъв случай не означава "прост". Напротив, клетката е уникално фрактално творение на Бог, поразително със своята сложност и в същото време с изключителната съгласуваност на работата на всички негови елементи.
Когато успяхме да надникнем вътре с помощта на електронен микроскоп, се оказа, че структурата на простата клетка е толкова сложна и неразбираема, колкото самата Вселена. Днес вече е установено, че „Една клетка е специална материя на Вселената, специална материя на Космоса“. Една единствена клетка съдържа информация, която може да бъде поставена само в няколко десетки хиляди тома на Голямата съветска енциклопедия. Тези. клетката, освен всичко друго, е огромен "биорезервоар" от информация.
Авторът на съвременната теория на молекулярната еволюция Манфред Айген пише: „За да се образува случайно протеинова молекула, природата ще трябва да направи около 10130 опита и да изразходва за това такъв брой молекули, които биха били достатъчни за 1027 Вселени. Ако протеинът е изграден интелигентно, тоест, че валидността на всеки ход може да бъде проверена чрез някакъв механизъм за селекция, са били необходими само около 2000 опита. Стигаме до парадоксален извод: програмата за изграждане на "примитивна жива клетка" е кодиран някъде на нивото на елементарните частици".
И как би могло да бъде иначе. Всяка клетка, притежаваща ДНК, е надарена със съзнание, осъзнава себе си и другите клетки и е в контакт с Вселената, като всъщност е част от нея. И въпреки че броят и разнообразието от клетки в човешкото тяло са невероятни (около 70 трилиона), всички те са себеподобни, точно както всички процеси, протичащи в клетките, са себеподобни. По думите на немския учен Роланд Глазер, дизайнът на биологичните клетки е „много добре обмислен“. Кой е добре обмислен?
Отговорът е прост: протеините, нуклеиновите киселини, живите клетки и всички биологични системи са продукт на творческата дейност на интелектуалния Създател.
Интересното е: на атомно ниво няма разлики между химическия състав на органичния и неорганичния свят. С други думи, на ниво атом клетката се създава от същите елементи като неживата природа. Разликите се откриват на молекулярно ниво. В живите тела, наред с неорганичните вещества и водата, има също протеини, въглехидрати, мазнини, нуклеинови киселини, ензима АТФ синтаза и други нискомолекулни органични съединения.
Към днешна дата клетката е буквално разглобена на атоми с цел изследване. Не е възможно обаче да се създаде поне една жива клетка, защото да създадеш клетка означава да създадеш частица от живата Вселена. Академик В.П. Казначеев смята, че „една клетка е космопланетен организъм... Човешките клетки са определени системи от ефирно-торсионни биоколайдери. В тези биоколайдери протичат непознати за нас процеси, осъществява се материализиране на космически форми на потоци, тяхното космическо преобразуване и поради това частиците се материализират".
Вода.
Почти 80% от клетъчната маса е вода. Според д-р по биология С. Зенин водата, поради своята клъстерна структура, е информационна матрица за управление на биохимичните процеси. Освен това водата е основната "мишена", с която взаимодействат колебанията на звуковата честота. Подредеността на клетъчната вода е толкова висока (близка до подредеността на кристала), че се нарича течен кристал.
катерици.
Протеините играят важна роля в биологичния живот. Клетката съдържа няколко хиляди протеини, които са уникални за този тип клетки (с изключение на стволовите клетки). Способността да синтезира собствени протеини се наследява от клетка на клетка и се запазва през целия живот. По време на живота на клетката протеините постепенно променят структурата си, функцията им се нарушава. Тези изразходвани протеини се отстраняват от клетката и се заменят с нови, благодарение на което се запазва жизнената активност на клетката.
Преди всичко отбелязваме строителната функция на протеините, тъй като те са строителният материал, който изгражда мембраните на клетките и клетъчните органели, стените на кръвоносните съдове, сухожилията, хрущялите и т.н.
Сигналната функция на протеините е изключително интересна. Оказва се, че протеините са в състояние да служат като сигнални вещества, предаващи сигнали между тъкани, клетки или организми. Сигналната функция се изпълнява от хормонални протеини. Клетките могат да комуникират помежду си на разстояние, използвайки сигнални протеини, предавани през междуклетъчното вещество.
Протеините имат и двигателна функция. Всички видове движения, на които клетките са способни, като мускулна контракция, се извършват от специални контрактилни протеини. Протеините изпълняват и транспортна функция. Те са в състояние да прикрепят различни вещества и да ги прехвърлят от едно място в клетката на друго. Например, кръвният протеин хемоглобин свързва кислород и го пренася до всички тъкани и органи на тялото. Освен това протеините имат и защитна функция. Когато в тялото се въвеждат чужди протеини или клетки, в него се произвеждат специални протеини, които свързват и неутрализират чужди клетки и вещества. И накрая, енергийната функция на протеините е, че при пълното разграждане на 1 g протеин се отделя енергия в размер на 17,6 kJ.
Клетъчна структура.
Клетката се състои от три неразривно свързани части: мембрана, цитоплазма и ядро, като структурата и функцията на ядрото в различните периоди от живота на клетката са различни. Животът на една клетка включва два периода: делене, в резултат на което се образуват две дъщерни клетки, и период между деленията, който се нарича интерфаза.
Клетъчната мембрана взаимодейства директно с външната среда и взаимодейства със съседните клетки. Състои се от външен слой и плазмена мембрана, разположена отдолу. Повърхностният слой на животинските клетки се нарича гликокали. Той свързва клетките с външната среда и с всички заобикалящи я вещества. Дебелината му е по-малка от 1 микрон.
Клетъчна структура
Клетъчната мембрана е много важна част от клетката. Той държи заедно всички клетъчни компоненти и ограничава външната и вътрешната среда.
Между клетките и външната среда има постоянен обмен на вещества. От външната среда в клетката влизат вода, различни соли под формата на отделни йони, неорганични и органични молекули. Продуктите на метаболизма, както и веществата, синтезирани в клетката: протеини, въглехидрати, хормони, които се произвеждат в клетките на различни жлези, се екскретират във външната среда през мембраната от клетката. Транспортирането на вещества е една от основните функции на плазмената мембрана.
Цитоплазма- вътрешна полутечна среда, в която протичат основните метаболитни процеси. Последните проучвания показват, че цитоплазмата не е вид разтвор, чиито компоненти взаимодействат помежду си при произволни сблъсъци. Може да се сравни с желето, което започва да "трепери" в отговор на външни влияния. Така цитоплазмата възприема и предава информация.
В цитоплазмата са разположени ядрото и различни органели, които са обединени от нея в едно цяло, което осигурява тяхното взаимодействие и дейността на клетката като единна интегрална система. Ядрото се намира в централната част на цитоплазмата. Цялата вътрешна зона на цитоплазмата е изпълнена с ендоплазмения ретикулум, който е клетъчен органоид: система от тубули, везикули и "цистерни", ограничени от мембрани. Ендоплазменият ретикулум участва в метаболитните процеси, осигурявайки транспорта на вещества от околната среда до цитоплазмата и между отделните вътреклетъчни структури, но основната му функция е участието в протеиновия синтез, който се осъществява в рибозомите. - микроскопични малки тела с кръгла форма с диаметър 15-20 nm. Синтезираните протеини първо се натрупват в каналите и кухините на ендоплазмения ретикулум и след това се транспортират до органелите и клетъчните места, където се консумират.
Освен протеини, цитоплазмата съдържа и митохондрии, малки тела с размери 0,2-7 микрона, които се наричат "електростанции" на клетките. Редокс реакциите протичат в митохондриите, осигурявайки клетките с енергия. Броят на митохондриите в една клетка варира от няколко до няколко хиляди.
Ядро- жизненоважната част на клетката, контролира синтеза на протеини и чрез тях всички физиологични процеси в клетката. В ядрото на неделяща се клетка се разграничават ядрената мембрана, ядреният сок, ядрото и хромозомите. Чрез ядрената обвивка се осъществява непрекъснат обмен на вещества между ядрото и цитоплазмата. Под ядрената обвивка - ядрен сок (полутечно вещество), който съдържа ядрото и хромозомите. Ядрото е плътно закръглено тяло, чиито размери могат да варират в широки граници, от 1 до 10 микрона и повече. Състои се основно от рибонуклеопротеини; участва в образуването на рибозоми. Обикновено в клетката има 1-3 нуклеоли, понякога до няколкостотин. Ядрото се състои от РНК и протеин.
С появата на клетката на Земята възниква Животът!
Следва продължение...


