უზრუნველყოფს უჯრედის ურთიერთობას გარემოსთან. ორგანიზმის კავშირი გარემოსთან
ორგანიზმის კავშირი გარემოსთან, ფიზიკურ-ქიმიური თვალსაზრისით, არის ღია სისტემა, ანუ სისტემა, სადაც ბიოქიმიური პროცესები მიმდინარეობს. საწყისი ნივთიერებები მოდის გარემოდან და ის ნივთიერებები, რომლებიც ასევე მუდმივად წარმოიქმნება, გამოიყოფა. ორგანიზმში მრავალმხრივი რეაქციების პროდუქტების სისწრაფესა და კონცენტრაციას შორის ბალანსი პირობითი, წარმოსახვითია, ვინაიდან ნივთიერებების მიღება და მოცილება არ ჩერდება. მუდმივი კავშირი გარემოსთან და გვაძლევს საშუალებას მივიჩნიოთ ცოცხალი ორგანიზმი, როგორც ღია სისტემა.
მზე არის ენერგიის წყარო ყველა ცოცხალი უჯრედისთვის. მცენარეული უჯრედები მზის სინათლის ენერგიას ქლოროფილის დახმარებით ითვისებენ, ფოტოსინთეზის დროს ასიმილაციის რეაქციებისათვის. ცხოველების, სოკოების, ბაქტერიების უჯრედები ირიბად იყენებენ მზის ენერგიას მიწიერი მცენარის მიერ სინთეზირებული ორგანული ნივთიერებების გაყოფისას.
უჯრედის საკვები ნივთიერებების ნაწილი იშლება უჯრედული სუნთქვის პროცესში, რითაც ამარაგებს სხვადასხვა სახის უჯრედული აქტივობისთვის საჭირო ენერგიას. ეს პროცესი ხდება ორგანელებში, რომელსაც მიტოქონდრია ეწოდება. მიტოქონდრიონი შედგება ორი მემბრანისგან: გარე, რომელიც გამოყოფს ორგანელას ციტოპლაზმისგან და შიდა, რომელიც ქმნის მრავალ ნაკეცს. სუნთქვის მთავარი პროდუქტი არის ATP. ის ტოვებს მიტოქონდრიას და გამოიყენება როგორც ენერგიის წყარო ციტოპლაზმაში და უჯრედის მემბრანაში მრავალი ქიმიური რეაქციისთვის. თუ უჯრედული სუნთქვის განსახორციელებლად საჭიროა ჟანგბადი, მაშინ სუნთქვას ეწოდება აერობული სუნთქვა, მაგრამ თუ რეაქციები ხდება ჟანგბადის არარსებობის პირობებში, მაშინ საუბარია ანაერობულ სუნთქვაზე.
უჯრედში შესრულებული ნებისმიერი სახის სამუშაოსთვის ენერგია გამოიყენება ერთი ფორმით - ატფ-ის ფოსფატური ბმებიდან მიღებული ენერგიის სახით. ATP არის ძალიან მობილური ნაერთი. ATP წარმოიქმნება მიტოქონდრიის შიდა მემბრანაზე. ATP სინთეზირდება ყველა უჯრედში სუნთქვის დროს ნახშირწყლების, ცხიმების და სხვა ორგანული ნივთიერებების დაჟანგვის ენერგიის გამო. მწვანე მცენარეების უჯრედებში ატფ-ის ძირითადი რაოდენობა სინთეზირდება ქლოროპლასტებში მზის ენერგიის გამო. მათში, ფოტოსინთეზის დროს, ბევრჯერ მეტი ATP წარმოიქმნება, ვიდრე მიტოქონდრიაში. ATP იშლება ფოსფორ-ჟანგბადის ბმების გაწყვეტით და ენერგიის გამოყოფით. ეს ხდება ATPase ფერმენტის მოქმედებით ATP ჰიდროლიზის პროცესში - წყლის დამატებით ფოსფორმჟავას მოლეკულის ელიმინაციასთან ერთად. შედეგად, ATP გარდაიქმნება ADP-ად და თუ ფოსფორის მჟავის ორი მოლეკულა იყოფა, მაშინ AMP-ად. მჟავის თითოეული გრამ-მოლეკულის დაშლის რეაქციას თან ახლავს 40 კჯ გამოყოფა. ეს არის ძალიან დიდი ენერგეტიკული გამოსავალი, ამიტომ ATP-ის ფოსფორ-ჟანგბადის ობლიგაციებს ჩვეულებრივ უწოდებენ მაკროერგიულ (მაღალ ენერგიას).
პლასტიკური გაცვლის რეაქციებში ატფ-ის გამოყენება ხორციელდება ატფ-ის ჰიდროლიზთან მათი კონიუგაციის გზით. სხვადასხვა ნივთიერების მოლეკულები ენერგიით დამუხტულია ATP მოლეკულიდან ჰიდროლიზის დროს გამოთავისუფლებული ფოსფორის ჯგუფის მიმაგრებით, ანუ ფოსფორილირებით.
ფოსფატის წარმოებულების თავისებურება ის არის, რომ ისინი ვერ ტოვებენ უჯრედს, თუმცა მათი „გამოშვებული“ ფორმები თავისუფლად გადის მემბრანაში. ამის გამო, ფოსფორილირებული მოლეკულები უჯრედში რჩება მანამ, სანამ არ გამოიყენებენ შესაბამის რეაქციებს.
ADP-ის ATP-ად გადაქცევის საპირისპირო პროცესი ხდება ADP-ზე ფოსფორმჟავას მოლეკულის მიმაგრებით, წყლის გათავისუფლებით და დიდი რაოდენობით ენერგიის შთანთქმით.
ამრიგად, ATP არის ენერგიის უნივერსალური და უშუალო წყარო უჯრედების აქტივობისთვის. ეს ქმნის ენერგიის ერთიან ფიჭურ ფონდს და შესაძლებელს ხდის მის გადანაწილებას და ტრანსპორტირებას უჯრედის ერთი ნაწილიდან მეორეზე.
ფოსფატის ჯგუფის გადაცემა მნიშვნელოვან როლს ასრულებს ქიმიურ რეაქციებში, როგორიცაა მონომერებიდან მაკრომოლეკულების შეკრება. მაგალითად, ამინომჟავები შეიძლება გაერთიანდეს მხოლოდ პეპტიდებში, თუ ისინი ადრე ფოსფორილირებულია. შეკუმშვის ან მოძრაობის მექანიკური პროცესები, ხსნარის გადატანა კონცენტრაციის გრადიენტთან მიმართებაში და სხვა პროცესები დაკავშირებულია ATP-ში შენახული ენერგიის ხარჯვასთან.
ენერგიის გაცვლის პროცესი შეიძლება წარმოდგენილი იყოს შემდეგნაირად. ციტოპლაზმაში მაღალმოლეკულური ორგანული ნივთიერებები ფერმენტულად, ჰიდროლიზით, გარდაიქმნება უფრო მარტივებად, რომელთაგან შედგება: ცილები - ამინომჟავებად, პოლი- და დისაქარიდები - მონოსაქარიდებად (+ გლუკოზა), ცხიმები გლიცეროლად და ცხიმოვან მჟავებად. არ არის ოქსიდაციური პროცესები, გამოიყოფა მცირე ენერგია, რომელიც არ გამოიყენება და გადადის თერმულ ფორმაში. უჯრედების უმეტესობა პირველ რიგში იყენებს ნახშირწყლებს. პოლისაქარიდები (სახამებელი მცენარეებში და გლიკოგენი ცხოველებში) ჰიდროლიზდება გლუკოზამდე. გლუკოზის დაჟანგვა ხდება სამ ფაზაში: გლიკოლიზი, ოქსიდაციური დეკარბოქსილაცია (კრების ციკლი - ლიმონმჟავას ციკლი) და ოქსიდაციური ფოსფორილირება (რესპირატორული ჯაჭვი). გლიკოლიზი, რომლის შედეგადაც გლუკოზის ერთი მოლეკულა იყოფა პირუვიკ მჟავას ორ მოლეკულად ატფ-ის ორი მოლეკულის გამოყოფით, ციტოპლაზმაში მიმდინარეობს. ჟანგბადის არარსებობის შემთხვევაში, პირუვიკის მჟავა გარდაიქმნება ეთანოლში (დუღილი) ან რძემჟავა (ანაერობული სუნთქვა).
თუ გლიკოლიზი ტარდება ცხოველურ უჯრედებში, ექვსნახშირბადოვანი გლუკოზის მოლეკულა იშლება რძემჟავას ორ მოლეკულად. ეს პროცესი მრავალსაფეხურიანია. იგი თანმიმდევრულად ხორციელდება 13 ფერმენტის მიერ. ალკოჰოლური დუღილის დროს გლუკოზის მოლეკულისგან წარმოიქმნება ეთანოლის ორი მოლეკულა და CO2-ის ორი მოლეკულა.
გლიკოლიზი არის ანაერობული და აერობული სუნთქვის საერთო ფაზა, დანარჩენი ორი ტარდება მხოლოდ აერობულ პირობებში. ჟანგბადისგან თავისუფალი დაჟანგვის პროცესი, რომლის დროსაც მეტაბოლიტების ენერგიის მხოლოდ ნაწილი გამოიყოფა და გამოიყენება, არის საბოლოო პროცესი ანაერობული ორგანიზმებისთვის. ჟანგბადის თანდასწრებით, პირუვიკ მჟავა გადადის მიტოქონდრიაში, სადაც, რიგი თანმიმდევრული რეაქციების შედეგად, იგი მთლიანად იჟანგება აერობული გზით H2O-მდე და CO2-მდე ADP-ის ერთდროული ფოსფორილირებით ATP-მდე. ამავდროულად, გლიკოლიზი იძლევა ორ ATP მოლეკულას, ორი - კრებსის ციკლს, 34 - რესპირატორულ ჯაჭვს. წმინდა გამოსავალი გლუკოზის ერთი მოლეკულის H2O და CO2-მდე სრული დაჟანგვიდან არის 38 მოლეკულა.
ამრიგად, აერობულ ორგანიზმებში ორგანული ნივთიერებების საბოლოო დაშლა ხდება ატმოსფერული ჟანგბადით მათი დაჟანგვით მარტივ არაორგანულ ნივთიერებებად: CO2 და H2O. ეს პროცესი მიტოქონდრიის კრისტაზე მიმდინარეობს. ამ შემთხვევაში გამოიყოფა თავისუფალი ენერგიის მაქსიმალური რაოდენობა, რომლის მნიშვნელოვანი ნაწილი რეზერვირებულია ATP მოლეკულებში. ადვილი მისახვედრია, რომ აერობული დაჟანგვა უზრუნველყოფს უჯრედს თავისუფალი ენერგიით უდიდეს ზომით.
კატაბოლიზმის შედეგად უჯრედში გროვდება ენერგიით მდიდარი ატფ-ის მოლეკულები და CO2 და ჭარბი წყალი გამოიყოფა გარე გარემოში.
შაქრის მოლეკულები, რომლებიც არ არის საჭირო სუნთქვისთვის, შეიძლება შეინახოს უჯრედში. ჭარბი ლიპიდები ან იშლება, რის შემდეგაც მათი დაშლის პროდუქტები შედიან მიტოქონდრიაში, როგორც სუნთქვის სუბსტრატი, ან დეპონირდება რეზერვში ციტოპლაზმაში ცხიმის წვეთების სახით. ცილები აგებულია უჯრედში შემავალი ამინომჟავებისგან. ცილის სინთეზი ხდება ორგანელებში, რომლებსაც რიბოსომები ეწოდება. თითოეული რიბოსომა შედგება ორი ქვენაწილაკისგან - დიდი და პატარა: ორივე ქვენაწილაკში შედის ცილის მოლეკულები და რნმ-ის მოლეკულები.
რიბოსომები ხშირად მიმაგრებულია მემბრანების სპეციალურ სისტემაზე, რომელიც შედგება ტანკებისა და ვეზიკულებისგან, ეგრეთ წოდებულ ენდოპლაზმურ რეტიკულუმზე (ER); უჯრედებში, რომლებიც აწარმოებენ უამრავ ცილას, ენდოპლაზმური ბადე ხშირად ძალიან კარგად არის განვითარებული და სავსეა რიბოზომებით. ზოგიერთი ფერმენტი ეფექტურია მხოლოდ იმ შემთხვევაში, თუ ისინი მიმაგრებულია მემბრანაზე. ლიპიდების სინთეზში მონაწილე ფერმენტების უმეტესობა აქ მდებარეობს. ამრიგად, ენდოპლაზმური რეტიკულუმი, როგორც ეს იყო, ერთგვარი უჯრედის დესკტოპია.
გარდა ამისა, ER ყოფს ციტოპლაზმას ცალკეულ განყოფილებებად, ანუ ნაწილებად, ანუ გამოყოფს ციტოპლაზმაში ერთდროულად მიმდინარე სხვადასხვა ქიმიურ პროცესებს და ამით ამცირებს ამ პროცესების ერთმანეთში ჩარევის ალბათობას.
ხშირად მოცემული უჯრედის მიერ წარმოქმნილი პროდუქტები გამოიყენება უჯრედის გარეთ. ასეთ შემთხვევებში, რიბოსომებზე სინთეზირებული ცილები გადის ენდოპლაზმური ბადის მემბრანებში და იფუთება მემბრანულ ვეზიკულებში, რომლებიც წარმოიქმნება მათ გარშემო, რომლებიც შემდეგ იჭრება ER-დან. ეს ბუშტები, ერთმანეთზე გაბრტყელებული და დაწყობილი, როგორც ბლინები გროვაში, ქმნიან დამახასიათებელ სტრუქტურას, რომელსაც ეწოდება გოლჯის კომპლექსი, ან გოლჯის აპარატი. გოლჯის აპარატში ყოფნის დროს ცილები გარკვეულ ცვლილებებს განიცდიან. როდესაც მათი უჯრედიდან გასვლის დრო დგება, მემბრანული ბუშტუკები უჯრედის მემბრანას ერწყმის და ცარიელდება, შიგთავსს გარეთ ასხამს, ანუ სეკრეცია ხდება ეგზოციტოზის გზით.
ლიზოსომები ასევე წარმოიქმნება გოლჯის აპარატში - საჭმლის მომნელებელი ფერმენტების შემცველი მემბრანული ტომრები. იმის გაგება, თუ როგორ აწარმოებს უჯრედი, აფუთავს და ექსპორტს ახორციელებს გარკვეულ ცილებს და როგორ „იცის“ რომელი ცილები უნდა შეინახოს თავისთვის, თანამედროვე ციტოლოგიის ერთ-ერთი ყველაზე მომხიბვლელი დარგია.
ნებისმიერი უჯრედის მემბრანა მუდმივად მოძრაობს და იცვლება. ER მემბრანები ნელა მოძრაობენ მთელ უჯრედში. ამ მემბრანების ცალკეული მონაკვეთები გამოყოფილია და ქმნის ვეზიკულებს, რომლებიც დროებით ხდება გოლჯის აპარატის ნაწილი და შემდეგ, ეგზოციტოზის პროცესში, ერწყმის უჯრედულ მემბრანას.
მოგვიანებით, მემბრანის მასალა ბრუნდება ციტოპლაზმაში, სადაც იგი ხელახლა გამოიყენება.
უჯრედში შემავალი ან მის მიერ გარედან გამოთავისუფლებული ნივთიერებების გაცვლა, აგრეთვე სხვადასხვა სიგნალების გაცვლა მიკრო და მაკროგარემოსთან, ხდება უჯრედის გარე მემბრანის მეშვეობით. როგორც ცნობილია, უჯრედის მემბრანა არის ლიპიდური ორშრე, რომელშიც ჩაშენებულია სხვადასხვა ცილის მოლეკულები, რომლებიც მოქმედებენ როგორც სპეციალიზებული რეცეპტორები, იონური არხები, მოწყობილობები, რომლებიც აქტიურად გადასცემენ ან აშორებენ სხვადასხვა ქიმიკატებს, უჯრედშორის კონტაქტებს და ა.შ. ჯანსაღ ევკარიოტულ უჯრედებში ფოსფოლიპიდები ნაწილდება მემბრანა ასიმეტრიულად: გარე ზედაპირი შედგება სფინგომიელინისა და ფოსფატიდილქოლინისგან, შიდა ზედაპირი შედგება ფოსფატიდილსერინისა და ფოსფატიდილეთანოლამინისგან. ასეთი ასიმეტრიის შესანარჩუნებლად საჭიროა ენერგიის ხარჯვა. ამიტომ უჯრედის დაზიანების, მისი ინფექციის, ენერგეტიკული შიმშილის შემთხვევაში მემბრანის გარე ზედაპირი მდიდრდება მისთვის უჩვეულო ფოსფოლიპიდებით, რაც ხდება სიგნალი სხვა უჯრედებისთვის და ფერმენტებისთვის უჯრედის დაზიანების შესახებ მასზე შესაბამისი რეაქციით. უმნიშვნელოვანეს როლს ასრულებს ფოსფოლიპაზა A2-ის ხსნადი ფორმა, რომელიც არღვევს არაქიდონის მჟავას და ქმნის ლიზოფორმებს ზემოაღნიშნული ფოსფოლიპიდებისგან. არაქიდონის მჟავა არის შემზღუდველი რგოლი ისეთი ანთებითი შუამავლების შესაქმნელად, როგორიცაა ეიკოსანოიდები, ხოლო დამცავი მოლეკულები - პენტრაქსინები (C-რეაქტიული ცილა (CRP), ამილოიდური ცილების წინამორბედები) - მიმაგრებულია მემბრანის ლიზოფორმებზე, რასაც მოჰყვება აქტივაცია. კომპლემენტის სისტემა კლასიკური გზის გასწვრივ და უჯრედების განადგურება.
მემბრანის სტრუქტურა ხელს უწყობს უჯრედის შიდა გარემოს მახასიათებლების შენარჩუნებას, მის განსხვავებებს გარე გარემოსგან. ამას უზრუნველყოფს უჯრედის მემბრანის შერჩევითი გამტარიანობა, მასში აქტიური სატრანსპორტო მექანიზმების არსებობა. მათი დარღვევა პირდაპირი დაზიანების შედეგად, მაგალითად, ტეტროდოტოქსინის, უაბაინის, ტეტრაეთილამონიუმის, ან შესაბამისი „ტუმბოების“ არასაკმარისი ენერგომომარაგების შემთხვევაში, იწვევს უჯრედის ელექტროლიტური შემადგენლობის დარღვევას, მისი მეტაბოლიზმის ცვლილებას. , სპეციფიკური ფუნქციების დარღვევა - შეკუმშვა, აგზნების იმპულსის გატარება და ა.შ. ადამიანებში უჯრედული იონური არხების (კალციუმი, ნატრიუმი, კალიუმი და ქლორიდი) დარღვევა ასევე შეიძლება გენეტიკურად განისაზღვროს ამ გენების სტრუქტურაზე პასუხისმგებელი მუტაციით. არხები. ეგრეთ წოდებული არხოპათია არის ნერვული, კუნთოვანი და საჭმლის მომნელებელი სისტემის მემკვიდრეობითი დაავადებების მიზეზი. უჯრედის შიგნით წყლის გადაჭარბებულმა მიღებამ შეიძლება გამოიწვიოს მისი რღვევა - ციტოლიზი - მემბრანის პერფორაციის გამო კომპლემენტის გააქტიურების ან ციტოტოქსიური ლიმფოციტების და ბუნებრივი მკვლელების შეტევის დროს.
უჯრედის მემბრანაში ჩაშენებულია მრავალი რეცეპტორი – სტრუქტურები, რომლებიც შესაბამის სპეციფიკურ სასიგნალო მოლეკულებთან (ლიგანდებთან) შერწყმისას სიგნალს გადასცემენ უჯრედში. ეს ხდება სხვადასხვა მარეგულირებელი კასკადების მეშვეობით, რომლებიც შედგება ფერმენტულად აქტიური მოლეკულებისგან, რომლებიც თანმიმდევრულად აქტიურდებიან და საბოლოოდ ხელს უწყობენ სხვადასხვა უჯრედული პროგრამების განხორციელებას, როგორიცაა ზრდა და გამრავლება, დიფერენციაცია, მოძრაობა, დაბერება და უჯრედების სიკვდილი. მარეგულირებელი კასკადები საკმაოდ მრავალრიცხოვანია, მაგრამ მათი რაოდენობა ჯერ ბოლომდე დადგენილი არ არის. რეცეპტორების სისტემა და მათთან დაკავშირებული მარეგულირებელი კასკადები ასევე არსებობს უჯრედის შიგნით; ისინი ქმნიან გარკვეულ მარეგულირებელ ქსელს კონცენტრაციის წერტილებით, განაწილებით და შემდგომი სიგნალის გზის არჩევით, რაც დამოკიდებულია უჯრედის ფუნქციურ მდგომარეობაზე, მისი განვითარების სტადიაზე და სხვა რეცეპტორების სიგნალების ერთდროულ მოქმედებაზე. ამის შედეგი შეიძლება იყოს სიგნალის დათრგუნვა ან გაძლიერება, მისი მიმართულება სხვადასხვა მარეგულირებელი გზის გასწვრივ. როგორც რეცეპტორების აპარატი, ასევე სიგნალის გადაცემის გზები მარეგულირებელ კასკადებში, მაგალითად, ბირთვში, შეიძლება დაირღვეს გენეტიკური დეფექტის შედეგად, რომელიც წარმოიქმნება როგორც თანდაყოლილი დეფექტი ორგანიზმის დონეზე ან სომატური მუტაციის გამო გარკვეულ უჯრედში. ტიპი. ეს მექანიზმები შეიძლება დაზიანდეს ინფექციური აგენტებით, ტოქსინებით და ასევე შეიცვალოს დაბერების დროს. ამის საბოლოო ეტაპი შეიძლება იყოს უჯრედის ფუნქციების დარღვევა, მისი გამრავლებისა და დიფერენცირების პროცესები.
მოლეკულები, რომლებიც მნიშვნელოვან როლს ასრულებენ უჯრედშორისი ურთიერთქმედების პროცესებში, ასევე განლაგებულია უჯრედების ზედაპირზე. ეს შეიძლება შეიცავდეს უჯრედის ადჰეზიურ პროტეინებს, ქსოვილებთან თავსებადობის ანტიგენებს, ქსოვილის სპეციფიკურ, დიფერენცირებელ ანტიგენებს და ა.შ. ამ მოლეკულების შემადგენლობის ცვლილება იწვევს უჯრედშორისი ურთიერთქმედების დარღვევას და შეიძლება გამოიწვიოს ასეთი უჯრედების ელიმინაციის შესაბამისი მექანიზმების გააქტიურება, რადგან ისინი გარკვეულ საფრთხეს უქმნიან სხეულის მთლიანობას, როგორც ინფექციის რეზერვუარს, განსაკუთრებით ვირუსულს, ან როგორც სიმსივნის ზრდის პოტენციურ ინიციატორებს.
უჯრედის ენერგომომარაგების დარღვევა
უჯრედში ენერგიის წყაროა საკვები, რომლის დაშლის შემდეგ ენერგია გამოიყოფა საბოლოო ნივთიერებებამდე. მიტოქონდრია არის ენერგიის წარმოების მთავარი ადგილი, რომელშიც ნივთიერებები იჟანგება რესპირატორული ჯაჭვის ფერმენტების დახმარებით. ოქსიდაცია არის ენერგიის მთავარი მიმწოდებელი, რადგან გლიკოლიზის შედეგად, ენერგიის არაუმეტეს 5% გამოიყოფა დაჟანგვის სუბსტრატების (გლუკოზა) იგივე რაოდენობით, დაჟანგვასთან შედარებით. დაჟანგვის დროს გამოთავისუფლებული ენერგიის დაახლოებით 60% გროვდება ოქსიდაციური ფოსფორილირებით მაკროენერგიულ ფოსფატებში (ATP, კრეატინ ფოსფატი), დანარჩენი გამოიყოფა სითბოს სახით. მომავალში, მაღალი ენერგიის ფოსფატებს უჯრედი იყენებს ისეთი პროცესებისთვის, როგორიცაა ტუმბო, სინთეზი, გაყოფა, მოძრაობა, სეკრეცია და ა.შ. არსებობს სამი მექანიზმი, რომელთა დაზიანებამ შეიძლება გამოიწვიოს უჯრედისთვის ენერგიის მიწოდების დარღვევა. : პირველი არის ენერგიის მეტაბოლიზმის ფერმენტების სინთეზის მექანიზმი, მეორე არის ჟანგვითი ფოსფორილირების მექანიზმი, მესამე - ენერგიის გამოყენების მექანიზმი.
მიტოქონდრიის სასუნთქ ჯაჭვში ელექტრონების ტრანსპორტირების დარღვევა ან ადფ-ის დაჟანგვისა და ფოსფორილირების გამოყოფა პროტონული პოტენციალის დაკარგვით - ATP წარმოქმნის მამოძრავებელი ძალა, იწვევს ჟანგვითი ფოსფორილირების შესუსტებას ისე, რომ ენერგიის უმეტესი ნაწილი იფანტება სითბოს ფორმა და მაკროერგიული ნაერთების რაოდენობა მცირდება. ადრენალინის გავლენის ქვეშ დაჟანგვისა და ფოსფორილირების განცალკევებას იყენებენ ჰომოიოთერმული ორგანიზმების უჯრედები სითბოს გამომუშავების გასაზრდელად გაციების დროს სხეულის მუდმივი ტემპერატურის შესანარჩუნებლად ან მისი მატების დროს ცხელების დროს. თირეოტოქსიკოზის დროს შეიმჩნევა მნიშვნელოვანი ცვლილებები მიტოქონდრიის სტრუქტურასა და ენერგეტიკულ ცვლაში. ეს ცვლილებები თავდაპირველად შექცევადია, მაგრამ გარკვეული პერიოდის შემდეგ ისინი შეუქცევადი ხდება: მიტოქონდრიის ფრაგმენტი, იშლება ან შეშუპება, კარგავს კრისტებს, გადაიქცევა ვაკუოლებად და საბოლოოდ აგროვებს ნივთიერებებს, როგორიცაა ჰიალინი, ფერიტინი, კალციუმი, ლიპოფუსცინი. სკორბუტის მქონე პაციენტებში მიტოქონდრია ერწყმის ქონდრიოსფეროს წარმოქმნას, შესაძლოა პეროქსიდის ნაერთებით მემბრანის დაზიანების გამო. მიტოქონდრიების მნიშვნელოვანი დაზიანება ხდება მაიონებელი გამოსხივების გავლენის ქვეშ, ნორმალური უჯრედის ავთვისებიანად გადაქცევის დროს.
მიტოქონდრია არის კალციუმის იონების მძლავრი საწყობი, სადაც მისი კონცენტრაცია ციტოპლაზმაში სიდიდის რამდენიმე რიგით მაღალია. როდესაც მიტოქონდრია დაზიანებულია, კალციუმი შედის ციტოპლაზმაში, რაც იწვევს პროტეინების გააქტიურებას უჯრედშიდა სტრუქტურების დაზიანებით და შესაბამისი უჯრედის ფუნქციების მოშლით, მაგალითად, კალციუმის კონტრაქტურებით ან თუნდაც "კალციუმის სიკვდილთან" ნეირონებში. მიტოქონდრიის ფუნქციური უნარის დარღვევის შედეგად მკვეთრად იზრდება თავისუფალი რადიკალების პეროქსიდის ნაერთების წარმოქმნა, რომლებსაც აქვთ ძალიან მაღალი რეაქტიულობა და შესაბამისად აზიანებენ უჯრედის მნიშვნელოვან კომპონენტებს - ნუკლეინის მჟავებს, ცილებს და ლიპიდებს. ეს ფენომენი შეინიშნება ეგრეთ წოდებული ოქსიდაციური სტრესის დროს და შეიძლება ჰქონდეს ნეგატიური შედეგები უჯრედის არსებობაზე. ამრიგად, გარეთა მიტოქონდრიული მემბრანის დაზიანებას თან ახლავს მემბრანთაშორის სივრცეში შემავალი ნივთიერებების ციტოპლაზმაში განთავისუფლება, უპირველეს ყოვლისა ციტოქრომ C და ზოგიერთი სხვა ბიოლოგიურად აქტიური ნივთიერება, რაც იწვევს ჯაჭვურ რეაქციებს, რომლებიც იწვევს უჯრედების დაპროგრამებულ სიკვდილს - აპოპტოზს. მიტოქონდრიული დნმ-ის დაზიანებით, თავისუფალი რადიკალების რეაქციები ამახინჯებს გენეტიკურ ინფორმაციას, რომელიც აუცილებელია გარკვეული რესპირატორული ჯაჭვის ფერმენტების ფორმირებისთვის, რომლებიც წარმოიქმნება სპეციალურად მიტოქონდრიებში. ეს იწვევს ჟანგვითი პროცესების კიდევ უფრო დიდ დარღვევას. მთლიანობაში, მიტოქონდრიის შინაგანი გენეტიკური აპარატი, ბირთვის გენეტიკურ აპარატთან შედარებით, ნაკლებად არის დაცული მავნე ზემოქმედებისგან, რომელსაც შეუძლია შეცვალოს მასში დაშიფრული გენეტიკური ინფორმაცია. შედეგად, მიტოქონდრიული დისფუნქცია ხდება მთელი ცხოვრების განმავლობაში, მაგალითად, დაბერების პროცესში, უჯრედის ავთვისებიანი ტრანსფორმაციის დროს, ასევე მემკვიდრეობითი მიტოქონდრიული დაავადებების ფონზე, რომლებიც დაკავშირებულია კვერცხუჯრედში მიტოქონდრიული დნმ-ის მუტაციასთან. ამჟამად აღწერილია 50-ზე მეტი მიტოქონდრიული მუტაცია, რომლებიც იწვევს ნერვული და კუნთოვანი სისტემების მემკვიდრეობით დეგენერაციულ დაავადებებს. ისინი ბავშვს გადაეცემა ექსკლუზიურად დედისგან, რადგან სპერმის მიტოქონდრია არ არის ზიგოტის ნაწილი და, შესაბამისად, ახალი ორგანიზმი.
გენეტიკური ინფორმაციის შენარჩუნებისა და გადაცემის დარღვევა
უჯრედის ბირთვი შეიცავს გენეტიკური ინფორმაციის უმეტეს ნაწილს და ამით უზრუნველყოფს მის ნორმალურ ფუნქციონირებას. სელექციური გენის ექსპრესიის დახმარებით კოორდინაციას უწევს უჯრედის მუშაობას ინტერფაზაში, ინახავს გენეტიკურ ინფორმაციას, ხელახლა ქმნის და გადასცემს გენეტიკურ მასალას უჯრედების გაყოფის პროცესში. დნმ-ის რეპლიკაცია და რნმ-ის ტრანსკრიფცია ხდება ბირთვში. სხვადასხვა პათოგენურმა ფაქტორებმა, როგორიცაა ულტრაიისფერი და მაიონებელი გამოსხივება, თავისუფალი რადიკალების დაჟანგვა, ქიმიკატები, ვირუსები, შეიძლება დააზიანოს დნმ. ვარაუდობენ, რომ თბილსისხლიანი ცხოველის თითოეული უჯრედი 1 დღეშია. კარგავს 10000-ზე მეტ ბაზას. ამას უნდა დაემატოს დაყოფის დროს კოპირებისას დარღვევები. თუ ეს დაზიანება გაგრძელდა, უჯრედი ვერ გადარჩებოდა. დაცვა მდგომარეობს მძლავრი სარემონტო სისტემების არსებობაში, როგორიცაა ულტრაიისფერი ენდონუკლეაზა, რეპარაციული რეპლიკაცია და რეკომბინაციული სარემონტო სისტემები, რომლებიც ცვლის დნმ-ის დაზიანებას. რეპარაციულ სისტემებში გენეტიკური დეფექტები იწვევს დაავადებების განვითარებას დნმ-ის დამაზიანებელი ფაქტორების მიმართ გაზრდილი მგრძნობელობის გამო. ეს არის პიგმენტური ქსეროდერმია, ისევე როგორც ზოგიერთი დაჩქარებული დაბერების სინდრომი, რომელსაც თან ახლავს ავთვისებიანი სიმსივნეების გაჩენის გაზრდილი ტენდენცია.
დნმ-ის რეპლიკაციის პროცესების რეგულირების სისტემა, მესინჯერი რნმ-ის (მრნმ) ტრანსკრიფცია, ნუკლეინის მჟავებიდან გენეტიკური ინფორმაციის გადათარგმნა ცილების სტრუქტურაში საკმაოდ რთული და მრავალდონიანია. გარდა მარეგულირებელი კასკადებისა, რომლებიც იწვევენ 3000-ზე მეტი ტრანსკრიფციის ფაქტორების მოქმედებას, რომლებიც ააქტიურებენ გარკვეულ გენებს, ასევე არსებობს მრავალდონიანი მარეგულირებელი სისტემა, რომელსაც შუამავლობს მცირე რნმ-ის მოლეკულები (ინტერფერენტული რნმ; RNAi). ადამიანის გენომი, რომელიც შედგება დაახლოებით 3 მილიარდი პურინისა და პირიმიდინის ფუძისგან, შეიცავს ცილის სინთეზზე პასუხისმგებელი სტრუქტურული გენების მხოლოდ 2%-ს. დანარჩენი უზრუნველყოფს მარეგულირებელი რნმ-ების სინთეზს, რომლებიც ტრანსკრიფციის ფაქტორებთან ერთად ააქტიურებენ ან ბლოკავს სტრუქტურული გენების მუშაობას დნმ-ის დონეზე ქრომოსომებში ან გავლენას ახდენს მესენჯერი რნმ-ის (მრნმ) ტრანსლაციის პროცესებზე პოლიპეპტიდური მოლეკულის ფორმირების დროს. ციტოპლაზმა. გენეტიკური ინფორმაციის დარღვევა შეიძლება მოხდეს როგორც სტრუქტურული გენების, ასევე დნმ-ის მარეგულირებელ ნაწილში შესაბამისი გამოვლინებით სხვადასხვა მემკვიდრეობითი დაავადებების სახით.
ბოლო დროს დიდი ყურადღება დაეთმო გენეტიკური მასალის ცვლილებებს, რომლებიც ხდება ორგანიზმის ინდივიდუალური განვითარების დროს და დაკავშირებულია დნმ-ის და ქრომოსომების გარკვეული მონაკვეთების ინჰიბირებასთან ან გააქტიურებასთან მათი მეთილაციის, აცეტილირების და ფოსფორილირების გამო. ეს ცვლილებები გრძელდება დიდი ხნის განმავლობაში, ზოგჯერ ორგანიზმის მთელი სიცოცხლის მანძილზე ემბრიოგენეზიდან სიბერემდე და ეპიგენომიურ მემკვიდრეობას უწოდებენ.
შეცვლილი გენეტიკური ინფორმაციის მქონე უჯრედების რეპროდუქციას ასევე ხელს უშლის მიტოზური ციკლის კონტროლის სისტემები (ფაქტორები). ისინი ურთიერთქმედებენ ციკლინდამოკიდებულ პროტეინ კინაზებთან და მათ კატალიზურ ქვედანაყოფებთან - ციკლინებთან - და ბლოკავენ უჯრედის მიერ სრული მიტოზური ციკლის გავლას, აჩერებენ გაყოფას პრესინთეტურ და სინთეზურ ფაზებს შორის საზღვარზე (ბლოკი G1/S) დნმ-ის შეკეთების დასრულებამდე. და თუ ეს შეუძლებელია, ისინი იწყებენ დაპროგრამებული სიკვდილის უჯრედებს. ამ ფაქტორებს მიეკუთვნება p53 გენი, რომლის მუტაცია იწვევს ტრანსფორმირებული უჯრედების პროლიფერაციაზე კონტროლის დაკარგვას; ის გვხვდება ადამიანის კიბოს შემთხვევების თითქმის 50%-ში. მიტოზური ციკლის გავლის მეორე საგუშაგო მდებარეობს G2/M საზღვარზე. აქ, ქრომოსომული მასალის სწორი განაწილება ქალიშვილ უჯრედებს შორის მიტოზის ან მეიოზის დროს კონტროლდება მექანიზმების კომპლექსის გამოყენებით, რომლებიც აკონტროლებენ უჯრედის ღეროს, ცენტრსა და ცენტრომერებს (კინეტოქორები). ამ მექანიზმების არაეფექტურობა იწვევს ქრომოსომების ან მათი ნაწილების განაწილების დარღვევას, რაც გამოიხატება რომელიმე ქალიშვილურ უჯრედში რაიმე ქრომოსომის არარსებობით (ანეუფლოიდია), დამატებითი ქრომოსომის არსებობით (პოლიპლოიდია), გამოყოფით. ქრომოსომის ნაწილი (წაშლა) და მისი გადატანა სხვა ქრომოსომაში (ტრანსლოკაცია). ასეთი პროცესები ძალიან ხშირად შეინიშნება ავთვისებიანი გადაგვარებული და ტრანსფორმირებული უჯრედების გამრავლების დროს. თუ ეს ხდება ჩანასახოვან უჯრედებთან მეიოზის დროს, ეს იწვევს ან ნაყოფის სიკვდილს ემბრიონის განვითარების ადრეულ ეტაპზე, ან ქრომოსომული დაავადების მქონე ორგანიზმის დაბადებას.
სიმსივნის ზრდის დროს უჯრედების უკონტროლო რეპროდუქცია ხდება იმ გენების მუტაციების შედეგად, რომლებიც აკონტროლებენ უჯრედების პროლიფერაციას და უწოდებენ ონკოგენებს. ამჟამად ცნობილ 70-ზე მეტ ონკოგენს შორის, მათი უმეტესობა წარმოადგენს უჯრედების ზრდის რეგულირების კომპონენტებს, ზოგიერთი არის ტრანსკრიფციის ფაქტორები, რომლებიც არეგულირებენ გენის აქტივობას, ასევე ფაქტორები, რომლებიც აფერხებენ უჯრედების გაყოფას და ზრდას. პროლიფერაციული უჯრედების გადაჭარბებული გაფართოების (გავრცელების) შემზღუდველი კიდევ ერთი ფაქტორია ქრომოსომების ბოლოების - ტელომერების დამოკლება, რომლებსაც არ შეუძლიათ სრულად გამრავლება წმინდა სტერული ურთიერთქმედების შედეგად, ამიტომ, თითოეული უჯრედის გაყოფის შემდეგ, ტელომერები მცირდება. ბაზების გარკვეული ნაწილი. ამრიგად, ზრდასრული ორგანიზმის მზარდი უჯრედები, გარკვეული რაოდენობის გაყოფის შემდეგ (ჩვეულებრივ, 20-დან 100-მდე, ორგანიზმის ტიპზე და მის ასაკზეა დამოკიდებული), ამოწურავს ტელომერის სიგრძეს და ჩერდება შემდგომი ქრომოსომის რეპლიკაცია. ეს ფენომენი არ ხდება სპერმატოგენურ ეპითელიუმში, ენტეროციტებსა და ემბრიონულ უჯრედებში ტელომერაზას ფერმენტის არსებობის გამო, რომელიც ყოველი გაყოფის შემდეგ აღადგენს ტელომერების სიგრძეს. ზრდასრული ორგანიზმების უმეტეს უჯრედებში ტელომერაზა იბლოკება, მაგრამ, სამწუხაროდ, ის აქტიურდება სიმსივნურ უჯრედებში.
ბირთვსა და ციტოპლაზმას შორის კავშირი, ნივთიერებების ტრანსპორტირება ორივე მიმართულებით ხორციელდება ფორების მეშვეობით ბირთვულ მემბრანაში ენერგიის მოხმარების სპეციალური სატრანსპორტო სისტემების მონაწილეობით. ამრიგად, ენერგია და პლასტიკური ნივთიერებები, სიგნალის მოლეკულები (ტრანსკრიფციის ფაქტორები) ტრანსპორტირდება ბირთვში. საპირისპირო ნაკადი ციტოპლაზმაში შემოაქვს mRNA-ს მოლეკულებს და გადასცემს რნმ-ს (tRNA), რიბოზომებს, რომლებიც აუცილებელია უჯრედში ცილის სინთეზისთვის. ნივთიერებების ტრანსპორტირების იგივე გზა თანდაყოლილია ვირუსებში, კერძოდ, როგორიცაა აივ. ისინი გადააქვთ თავიანთ გენეტიკურ მასალას მასპინძელი უჯრედის ბირთვში მისი შემდგომი ჩართვით მასპინძლის გენომში და ახლად წარმოქმნილი ვირუსული რნმ-ის გადატანით ციტოპლაზმაში ახალი ვირუსული ნაწილაკების შემდგომი ცილის სინთეზისთვის.
სინთეზის პროცესების დარღვევა
ცილის სინთეზის პროცესები ხდება ენდოპლაზმური ბადის ცისტერნებში, რომლებიც მჭიდრო კავშირშია ბირთვული მემბრანის ფორებთან, რომლის მეშვეობითაც რიბოსომები, tRNA და mRNA შედიან ენდოპლაზმურ რეტიკულუმში. აქ ხდება პოლიპეპტიდური ჯაჭვების სინთეზი, რომლებიც შემდგომში იძენენ საბოლოო ფორმას აგრანულ ენდოპლაზმურ რეტიკულუმში და ლამელარულ კომპლექსში (გოლგის კომპლექსი), სადაც ისინი განიცდიან პოსტტრანსლაციურ მოდიფიკაციას და ასოციაციას ნახშირწყლებისა და ლიპიდების მოლეკულებთან. ახლად წარმოქმნილი ცილის მოლეკულები არ რჩებიან სინთეზის ადგილზე, არამედ რთული რეგულირებადი პროცესის დახმარებით, რომელიც ე.წ. პროტეინის კინეზი, აქტიურად გადადიან უჯრედის იმ იზოლირებულ ნაწილში, სადაც ისინი შეასრულებენ დანიშნულ ფუნქციას. ამ შემთხვევაში, ძალიან მნიშვნელოვანი ნაბიჯია გადაცემული მოლეკულის სტრუქტურირება შესაბამის სივრცულ კონფიგურაციაში, რომელსაც შეუძლია შეასრულოს მისი თანდაყოლილი ფუნქცია. ასეთი სტრუქტურირება ხდება სპეციალური ფერმენტების დახმარებით ან სპეციალიზებული ცილის მოლეკულების მატრიცაზე - ჩაპერონები, რომლებიც ეხმარება ცილის მოლეკულას, ახლად წარმოქმნილ ან შეცვლილ გარე გავლენის გამო, შეიძინოს სწორი სამგანზომილებიანი სტრუქტურა. უჯრედზე არასასურველი ზემოქმედების შემთხვევაში, როდესაც არსებობს ცილის მოლეკულების სტრუქტურის დარღვევის შესაძლებლობა (მაგალითად, სხეულის ტემპერატურის მატებით, ინფექციური პროცესით, ინტოქსიკაციით), უჯრედში ჩაპერონების კონცენტრაცია. მკვეთრად იზრდება. ამიტომ, ასეთ მოლეკულებს ასევე უწოდებენ სტრესის ცილები, ან სითბოს შოკის ცილები. ცილის მოლეკულის სტრუქტურის დარღვევა იწვევს ქიმიურად ინერტული კონგლომერატების წარმოქმნას, რომლებიც დეპონირდება უჯრედში ან მის გარეთ ამილოიდოზის, ალცჰეიმერის დაავადების და ა.შ. ზოგჯერ წინასწარ სტრუქტურირებული ანალოგიური მოლეკულა შეიძლება იყოს მატრიცა და ამ შემთხვევაში, თუ პირველადი სტრუქტურა არ მოხდა სწორად, ყველა შემდგომი მოლეკულა ასევე იქნება დეფექტური. ეს ვითარება ხდება ეგრეთ წოდებულ პრიონის დაავადებებში (სკრაპი ცხვარში, ცოფი ძროხებში, კურუ, კრეიტცფელდტ-იაკობის დაავადება ადამიანებში), როდესაც ნერვული უჯრედის ერთ-ერთი მემბრანის ცილის დეფექტი იწვევს შიგნით ინერტული მასების შემდგომ დაგროვებას. უჯრედი და მისი სასიცოცხლო აქტივობის დარღვევა.
უჯრედში სინთეზის პროცესების დარღვევა შეიძლება მოხდეს მის სხვადასხვა ეტაპზე: რნმ-ის ტრანსკრიფცია ბირთვში, პოლიპეპტიდების ტრანსლაცია რიბოსომებში, პოსტტრანსლაციური მოდიფიკაცია, კრემისფერი მოლეკულის ჰიპერმეთილაცია და გლიკოზილაცია, ცილების ტრანსპორტირება და განაწილება უჯრედში და მათი მოცილება. გარედან. ამ შემთხვევაში შეიძლება დაფიქსირდეს რიბოსომების რაოდენობის ზრდა ან შემცირება, პოლირიბოსომების დაშლა, მარცვლოვანი ენდოპლაზმური ბადის ცისტერნების გაფართოება, მის მიერ რიბოზომების დაკარგვა, ვეზიკულების და ვაკუოლების წარმოქმნა. ასე რომ, ფერმკრთალი გომბეშოთი მოწამვლის შემთხვევაში ზიანდება რნმ პოლიმერაზას ფერმენტი, რაც არღვევს ტრანსკრიფციას. დიფტერიის ტოქსინი, რომელიც ააქტიურებს დრეკადობის ფაქტორს, არღვევს ტრანსლაციის პროცესებს, რაც იწვევს მიოკარდიუმის დაზიანებას. ზოგიერთი სპეციფიკური ცილის მოლეკულის სინთეზის დარღვევის მიზეზი შეიძლება იყოს ინფექციური აგენტები. მაგალითად, ჰერპესვირუსები აფერხებენ MHC ანტიგენის მოლეკულების სინთეზს და გამოხატვას, რაც მათ საშუალებას აძლევს ნაწილობრივ აიცილონ იმუნური კონტროლი, ხოლო ჭირის ბაცილები აფერხებენ მწვავე ანთების შუამავლების სინთეზს. უჩვეულო ცილების გამოჩენამ შეიძლება შეაჩეროს მათი შემდგომი დაშლა და გამოიწვიოს ინერტული ან თუნდაც ტოქსიკური მასალის დაგროვება. გარკვეულწილად, დაშლის პროცესების დარღვევამ ასევე შეიძლება ხელი შეუწყოს ამას.
დაშლის პროცესების დარღვევა
უჯრედში ცილის სინთეზის პარალელურად, მისი დაშლა მუდმივად ხდება. ნორმალურ პირობებში ამას აქვს მნიშვნელოვანი მარეგულირებელი და ფორმირების მნიშვნელობა, მაგალითად, ფერმენტების, ცილოვანი ჰორმონების და მიტოზური ციკლის ცილების არააქტიური ფორმების გააქტიურებისას. უჯრედების ნორმალური ზრდა და განვითარება მოითხოვს კარგად კონტროლირებულ ბალანსს ცილების და ორგანელების სინთეზსა და დეგრადაციას შორის. ამასთან, ცილის სინთეზის პროცესში, სინთეზური აპარატის მუშაობაში არსებული შეცდომების, ცილის მოლეკულის არანორმალური სტრუქტურის, ქიმიური და ბაქტერიული აგენტების მიერ მისი დაზიანების გამო, მუდმივად იქმნება დეფექტური მოლეკულების საკმაოდ დიდი რაოდენობა. ზოგიერთი შეფასებით, მათი წილი ყველა სინთეზირებული ცილების დაახლოებით მესამედია.
ძუძუმწოვრების უჯრედებს აქვთ რამდენიმე ძირითადი ცილის დეგრადაციის გზები:ლიზოსომური პროტეაზების (პენტიდური ჰიდროლაზების), კალციუმდამოკიდებული პროტეინაზების (ენდოპეპტიდაზების) და პროტეაზომის სისტემის მეშვეობით. გარდა ამისა, ასევე არსებობს სპეციალიზებული პროტეინაზები, როგორიცაა კასპაზები. მთავარი ორგანელა, რომელშიც ხდება ნივთიერებების დეგრადაცია ევკარიოტულ უჯრედებში, არის ლიზოსომა, რომელიც შეიცავს უამრავ ჰიდროლიზურ ფერმენტს. ენდოციტოზის და სხვადასხვა ტიპის აუტოფაგიის გამო ლიზოსომებსა და ფაგოლიზოსომებში განადგურებულია როგორც დეფექტური ცილის მოლეკულები, ასევე მთელი ორგანელები: დაზიანებული მიტოქონდრია, პლაზმური მემბრანის მონაკვეთები, ზოგიერთი უჯრედგარე ცილა, სეკრეტორული გრანულების შიგთავსი.
ცილის დეგრადაციის მნიშვნელოვანი მექანიზმია პროტეასომა, რთული მულტიკატალიზური პროტეინაზას სტრუქტურა, რომელიც ლოკალიზებულია ციტოზოლში, ბირთვში, ენდოპლაზმურ რეტიკულუმში და უჯრედის მემბრანაზე. ეს ფერმენტული სისტემა პასუხისმგებელია დაზიანებული ცილების დაშლაზე, ისევე როგორც ჯანსაღი ცილები, რომლებიც უნდა მოიხსნას უჯრედების ნორმალური ფუნქციონირებისთვის. ამ შემთხვევაში, განადგურებული ცილები წინასწარ შერწყმულია სპეციფიკურ უბიკიტინის პოლიპეპტიდთან. თუმცა, არაუბიკვიტირებული ცილები ასევე შეიძლება ნაწილობრივ განადგურდეს პროტეასომებში. პროტეაზომებში ცილის მოლეკულის დაშლა მოკლე პოლიპეპტიდებად (დამუშავება) მათი შემდგომი პრეზენტაციით MHC I ტიპის მოლეკულებთან ერთად მნიშვნელოვანი რგოლია ორგანიზმის ანტიგენური ჰომეოსტაზის იმუნური კონტროლის განხორციელებაში. პროტეაზომის ფუნქციის შესუსტებისას ხდება დაზიანებული და არასაჭირო ცილების დაგროვება, რაც თან ახლავს უჯრედების დაბერებას. ციკლინდამოკიდებული ცილების დეგრადაციის დარღვევა იწვევს უჯრედების გაყოფის დარღვევას, სეკრეტორული ცილების დეგრადაციას - ცისტოფიბროზის განვითარებას. პირიქით, პროტეასომების ფუნქციის მატება თან ახლავს ორგანიზმის დაქვეითებას (შიდსი, კიბო).
ცილების დეგრადაციის გენეტიკურად განსაზღვრული დარღვევით ორგანიზმი სიცოცხლისუნარიანი არ არის და ემბრიოგენეზის ადრეულ სტადიაზე კვდება. თუ ცხიმების ან ნახშირწყლების დაშლა ირღვევა, მაშინ ხდება დაგროვების დაავადებები (თეზაურიზმები). ამავდროულად, უჯრედის შიგნით გროვდება გარკვეული ნივთიერებების ან მათი არასრული დაშლის პროდუქტების ჭარბი რაოდენობა - ლიპიდები, პოლისაქარიდები, რაც მნიშვნელოვნად აზიანებს უჯრედის ფუნქციას. ყველაზე ხშირად ის შეინიშნება ღვიძლის ეპითელიოციტებში (ჰეპატოციტები), ნეირონებში, ფიბრობლასტებსა და მაკროფაგოციტებში.
ნივთიერებების დაშლის პროცესებში შეძენილი დარღვევები შეიძლება მოხდეს პათოლოგიური პროცესების შედეგად (მაგალითად, ცილების, ცხიმების, ნახშირწყლების და პიგმენტური დისტროფია) და თან ახლდეს უჩვეულო ნივთიერებების წარმოქმნა. ლიზოსომური პროტეოლიზის სისტემაში დარღვევები იწვევს შიმშილის დროს ადაპტაციის დაქვეითებას ან დატვირთვის გაზრდას, ზოგიერთი ენდოკრინული დისფუნქციის წარმოქმნას - ინსულინის, თირეოგლობულინის, ციტოკინების და მათი რეცეპტორების დონის დაქვეითებას. ცილის დეგრადაციის დარღვევა ანელებს ჭრილობის შეხორცების ტემპს, იწვევს ათეროსკლეროზის განვითარებას და გავლენას ახდენს იმუნურ პასუხზე. ჰიპოქსიის პირობებში, უჯრედშიდა pH-ის ცვლილებები, რადიაციული დაზიანება, ხასიათდება მემბრანის ლიპიდების გაზრდილი პეროქსიდაციის, აგრეთვე ლიზოსომოტროპული ნივთიერებების გავლენით - ბაქტერიული ენდოტოქსინები, ტოქსიკური სოკოების მეტაბოლიტები (სპოროფუზარინი), სილიციუმის ოქსიდის კრისტალები - ლიზოსომის მემბრანის სტაბილურობა. ცვლილებები, გააქტიურებული ლიზოსომური ფერმენტები გამოიყოფა ციტოპლაზმაში, რაც იწვევს უჯრედის სტრუქტურების განადგურებას და მის სიკვდილს.
Თავი 1
უჯრედის ფიზიოლოგიის საფუძვლები
ი.დუდელი
პლაზმური მემბრანა . ცხოველური უჯრედები შემოიფარგლება პლაზმური მემბრანით (ნახ. 1.1). მის სტრუქტურაზე, რომელიც ძალიან ჰგავს მრავალი უჯრედშიდა მემბრანის აგებულებას, უფრო დეტალურად შევეხებით. მემბრანის ძირითადი მატრიცა შედგება ლიპიდებიძირითადად ფოსფატიდილქოლინი. ეს ლიპიდები შედგება ჰიდროფილური სათავე ჯგუფისგან, რომელსაც გრძელი ჰიდროფობიური ნახშირწყალბადის ჯაჭვები აქვს მიმაგრებული. წყალში, ასეთი ლიპიდები სპონტანურად ქმნიან ორ ფენას 4-5 ნმ სისქის ფენას, რომელშიც ჰიდროფილური ჯგუფები წყლიან გარემოს აწყდებიან, ხოლო ჰიდროფობიური ნახშირწყალბადის ჯაჭვები განლაგებულია ორ რიგში, რომლებიც ქმნიან უწყლო ლიპიდურ ფაზას. უჯრედის მემბრანები წარმოადგენს ამ ტიპის ლიპიდურ ორ ფენას და შეიცავს გლიკოლიპიდებს, ქოლესტერინს და ფოსფოლიპიდებს (ნახ. 1.2). გლიკოლიპიდების ჰიდროფილური ნაწილი წარმოიქმნება ოლიგოსაქარიდებით. გლიკოლიპიდები ყოველთვის განლაგებულია პლაზმური მემბრანის გარე ზედაპირზე და მოლეკულის ოლიგოსაქარიდული ნაწილი გარემოში ჩაძირული თმავით არის ორიენტირებული. ფოსფოლიპიდებს შორის მიმოფანტული თითქმის თანაბარი რაოდენობით ქოლესტერინის მოლეკულები ასტაბილურებს მემბრანას. მემბრანის შიდა და გარე შრეებში სხვადასხვა ლიპიდების განაწილება ერთნაირი არ არის და ერთი და იგივე ფენის შიგნითაც არის ადგილები, რომლებშიც კონცენტრირებულია გარკვეული ტიპის ლიპიდები. ასეთი არათანაბარი განაწილება
ბრინჯი. 1.1. უჯრედის სქემატური ნახაზი, რომელიც აჩვენებს ყველაზე მნიშვნელოვან ორგანელებს
ალბათ აქვს გარკვეული, ჯერ კიდევ ბუნდოვანი, ფუნქციონალური მნიშვნელობა.
მემბრანის შედარებით ინერტული ლიპიდური მატრიცაში ჩაძირული ძირითადი ფუნქციური ელემენტებია ციყვები(ნახ. 1.2). პროტეინი წონით მერყეობს 25-დან 75%-მდე სხვადასხვა მემბრანაში, მაგრამ რადგან ცილის მოლეკულები ბევრად აღემატება ლიპიდურ მოლეკულებს, წონის 50% უდრის 1 ცილის მოლეკულის თანაფარდობას 50 ლიპიდურ მოლეკულასთან. ზოგიერთი ცილა აღწევს მემბრანაში მისი გარედან შიდა ზედაპირზე, ზოგი კი ფიქსირდება ერთ ფენაში. ცილის მოლეკულები, როგორც წესი, ისეა ორიენტირებული, რომ მათი ჰიდროფობიური ჯგუფები ჩაეფლო ლიპიდურ მემბრანაში და პოლარული ჰიდროფილური ჯგუფები მემბრანის ზედაპირზე ჩაეფლო წყალ ფაზაში. ბევრი გარე ზედაპირის მემბრანის ცილა არის გლიკოპროტეინი; მათი ჰიდროფილური საქარიდული ჯგუფები ხვდებიან უჯრედგარე გარემოს.
უჯრედშიდა ორგანელების მემბრანული სისტემები .
უჯრედის მოცულობის დაახლოებით ნახევარს იკავებს ორგანელები, რომლებიც იზოლირებულია ციტოზოლიდან მემბრანებით. უჯრედშიდა ორგანილების მემბრანების მთლიანი ზედაპირი მინიმუმ 10-ჯერ აღემატება პლაზმური მემბრანის ზედაპირს. ყველაზე ფართოდ გამოყენებული მემბრანული სისტემაა ენდოპლაზმურ ბადეში,წარმოადგენს ქსელს

ბრინჯი. 1.2.პლაზმური მემბრანის სქემატური წარმოდგენა. პროტეინები ჩაშენებულია ფოსფოლიპიდურ ორ შრეში, ზოგი მოიცავს ორ შრეს, ზოგი კი მხოლოდ გარე ან შიდა შრეზეა მიმაგრებული.
უაღრესად ჩახლართული მილაკები ან სახსრის წაგრძელებული სტრუქტურები; ენდოპლაზმური ბადის დიდი უბნები მოფენილია რიბოზომებით; ასეთ რეტიკულუმს მარცვლოვანი, ან უხეში ეწოდება (ნახ. 1.1). გოლგის აპარატიასევე შედგება გარსით შეკრული ლამელებისგან, საიდანაც იშლება ვეზიკულები, ანუ ვეზიკულები (ნახ. 1.1). ლიზოსომები და პეროქსიზომებიარის პატარა სპეციალიზებული ვეზიკულები. ყველა ამ მრავალფეროვან ორგანელებში მემბრანა და მას ფარავს სივრცე შეიცავს ფერმენტების სპეციფიკურ კომპლექტს; ორგანელების შიგნით გროვდება სპეციალური მეტაბოლური პროდუქტები, რომლებიც გამოიყენება ორგანელების სხვადასხვა ფუნქციების შესასრულებლად.
ბირთვიდა მიტოქონდრიაგანსხვავდება იმით, რომ თითოეული ეს ორგანელა გარშემორტყმულია ორი გარსით. ბირთვი პასუხისმგებელია მეტაბოლიზმის კინეტიკურ კონტროლზე; დაკეცილი შიდა მიტოქონდრიული მემბრანა არის ჟანგვითი მეტაბოლიზმის ადგილი; აქ, პირუვატის ან ცხიმოვანი მჟავების დაჟანგვის გამო, სინთეზირებულია მაღალი ენერგიის ნაერთი ადენოზინტრიფოსფატი (ATP, ან ATP).
ციტოჩონჩხი . ორგანელების მიმდებარე ციტოპლაზმა არავითარ შემთხვევაში არ შეიძლება ჩაითვალოს ამორფულად; იგი გაჟღენთილია ციტოჩონჩხის ქსელით. ციტოჩონჩხი შედგება მიკროტუბულებისგან, აქტინის ძაფებისგან და შუალედური ძაფებისგან (ნახ. 1.1). მიკროტუბულებიაქვს გარე დიამეტრი დაახლოებით 25 ნმ; ისინი წარმოიქმნება, როგორც ჩვეულებრივი პოლიმერი, ტუბულინის ცილის მოლეკულების შეკრების შედეგად. აქტინის ძაფები -შეკუმშვადი ბოჭკოები, რომლებიც მდებარეობს მემბრანულ შრეში და მთელ უჯრედში - ძირითადად მონაწილეობენ მოძრაობასთან დაკავშირებულ პროცესებში. შუალედური ძაფებიშედგება სხვადასხვა ტიპის უჯრედებში სხვადასხვა ქიმიური შემადგენლობის ბლოკებისგან; ისინი ქმნიან მრავალფეროვან კავშირებს ზემოთ ნახსენები ციტოჩონჩხის სხვა ორ ელემენტს შორის. ორგანელები და პლაზმური მემბრანა ასევე დაკავშირებულია ციტოჩონჩხთან, რომელიც არა მხოლოდ ინარჩუნებს უჯრედის ფორმას და მასში ორგანილების პოზიციას, არამედ განსაზღვრავს უჯრედის ფორმის ცვლილებას და მის მობილურობას.
ციტოზოლი . უჯრედის მოცულობის დაახლოებით ნახევარი იკავებს ციტოზოლს. იმის გამო, რომ ეს არის დაახლოებით 20% (წონის) პროტეინი, ის უფრო გელია, ვიდრე წყალხსნარი. მცირე მოლეკულები, მათ შორის ორგანული და არაორგანული იონები,იხსნება წყლის ფაზაში. უჯრედსა და გარემოს (უჯრედგარე სივრცე) შორის ხდება იონების გაცვლა; ეს გაცვლითი პროცესები განხილული იქნება შემდეგ ნაწილში. უჯრედგარე სივრცეში იონების კონცენტრაცია შენარჩუნებულია მნიშვნელოვანი სიზუსტით მუდმივ დონეზე; თითოეული იონის უჯრედშიდა კონცენტრაციას ასევე აქვს სპეციფიკური დონე, რომელიც განსხვავდება უჯრედის გარეთ (ცხრილი 1.1). უჯრედგარე გარემოში ყველაზე გავრცელებული კატიონია Na+ უჯრედში მისი კონცენტრაცია 10-ჯერ ნაკლებია. პირიქით, უჯრედის შიგნით K + კონცენტრაცია ყველაზე მაღალია, უჯრედის გარეთ ის უფრო დაბალია, ვიდრე სიდიდის ბრძანება. ყველაზე დიდი გრადიენტი უჯრედგარე და უჯრედშიდა კონცენტრაციებს შორის არსებობს Ca 2+-ისთვის, რომლის თავისუფალი იონების კონცენტრაცია უჯრედის შიგნით მინიმუმ 10000-ჯერ დაბალია, ვიდრე მის გარეთ. ყველა იონი არ იხსნება ციტოზოლში; ზოგიერთი მათგანი ადსორბირდება ცილებზე ან დეპონირდება ორგანელებში. მაგალითად, Ca 2+-ის შემთხვევაში შეკრული იონები ბევრად უფრო მრავალრიცხოვანია ვიდრე თავისუფალი. ციტოზოლის ცილების უმეტესობა ფერმენტებია, რომელთა მონაწილეობით ტარდება შუალედური მეტაბოლიზმის მრავალი პროცესი: გლიკოლიზი და გლუკონეოგენეზი, ამინომჟავების სინთეზი ან განადგურება, ცილის სინთეზი რიბოზომებზე (ნახ. 1.1). ციტოზოლი ასევე შეიცავს ლიპიდურ წვეთებს და გლიკოგენის გრანულებს, რომლებიც ემსახურებიან მნიშვნელოვანი მოლეკულების რეზერვებს.
ცხრილი 1.1.ჰომოიოთერმული ცხოველების კუნთოვან უჯრედებში იონების შიდა და უჯრედგარე კონცენტრაციები. მაგრამ – - "მაღალმოლეკულური წონის ფიჭური ანიონები"
|
უჯრედშიდა კონცენტრაცია |
უჯრედგარე კონცენტრაცია |
||
|
Na+ |
12 მმოლ/ლ |
Na+ |
145 მმოლ/ლ |
|
155 მმოლ/ლ |
K+ |
4 მმოლ/ლ |
|
|
Ca 2+ |
10 –7 10 –8 მმოლ/ლ |
Ca 2+ |
2 მმოლ/ლ |
|
ლ-თან ერთად - |
4 მმოლ/ლ |
C1 - |
120 მმოლ/ლ |
|
HCO 3 - |
8 მმოლ/ლ |
HCO 3 - |
27 მმოლ/ლ |
|
A- |
155 მმოლ/ლ |
სხვა კათიონები |
5 მმოლ/ლ |
|
მოსვენების პოტენციალი -90 მვ |
|||
1.2. ნივთიერებების გაცვლა უჯრედსა და გარემოს შორის
ჩვენ მოკლედ აღვწერეთ უჯრედის სტრუქტურა, რათა გამოვიყენოთ ეს აღწერილობა უჯრედის ფიზიოლოგიის საფუძვლების გადასახედად. უჯრედი არავითარ შემთხვევაში არ შეიძლება ჩაითვალოს სტატიკურ წარმონაქმნად, რადგან ხდება ნივთიერებების მუდმივი გაცვლა სხვადასხვა უჯრედშიდა ნაწილებს შორის, ასევე განყოფილებებსა და გარემოს შორის. უჯრედის სტრუქტურები დინამიურ წონასწორობაშია და უჯრედების ურთიერთქმედება ერთმანეთთან და გარე გარემოსთან აუცილებელი პირობაა მოქმედი ორგანიზმის სიცოცხლის შესანარჩუნებლად. ამ თავში განვიხილავთ ასეთი გაცვლის ფუნდამენტურ მექანიზმებს. შემდგომ თავებში ეს მექანიზმები განხილული იქნება ნერვულ უჯრედთან და მის ფუნქციებთან მიმართებაში;
თუმცა, იგივე მექანიზმები საფუძვლად უდევს ყველა სხვა ორგანოს ფუნქციონირებას.
დიფუზია.ნივთიერების გადაადგილების უმარტივესი პროცესია დიფუზია. ხსნარებში (ან აირებში) ატომები და მოლეკულები თავისუფლად მოძრაობენ და კონცენტრაციების სხვაობა დაბალანსებულია დიფუზიით. განვიხილოთ სითხით ან აირით სავსე ორი ტომი (ნახ. 1.3), რომლებშიც ნივთიერებებს აქვთ კონცენტრაცია c1 და c2 და გამოყოფილია ფენით A ზედაპირის ფართობით და სისქითდ. მატერიის დინება m დროში t აღწერილი ფიკის დიფუზიის პირველი კანონი:
დმ/ dt= და/ დ ( C 1 –С 2)=და/ დდ C(1)
სადაც D არის დიფუზიის კოეფიციენტი, რომელიც მუდმივია მოცემული ნივთიერებისთვის, გამხსნელისთვის და ტემპერატურისთვის. უფრო ზოგადი ფორმით, კონცენტრაციის სხვაობისთვის dc მანძილზე dx
dm/dt= -D A dc/dx,(2)
ნაკადი A მონაკვეთში კონცენტრაციის გრადიენტის პროპორციულია dc/dx . მინუს ნიშანი ჩნდება განტოლებაში, რადგან კონცენტრაციის ცვლილება x მიმართულებით უარყოფითია.
დიფუზია არის ყველაზე მნიშვნელოვანი პროცესი, რომლითაც წყალხსნარებში მოლეკულების უმეტესობა მოძრაობს მცირე მანძილზე. ეს ასევე ეხება მათ მოძრაობას უჯრედში, რამდენადაც დიფუზია არ არის შეფერხებული მემბრანებით. ბევრ ნივთიერებას შეუძლია თავისუფლად გავრცელდეს ლიპიდურ მემბრანებში, განსაკუთრებით წყალში და გახსნილ გაზებში, როგორიცაა O 2 და CO 2 . ცხიმში ხსნადი

ბრინჯი. 1.3.დიფუზიის რაოდენობრივი სქემა. ორი სივრცე გამოყოფილია სისქის ფენითდდა ფართობი მაგრამ. C; - ნაწილაკების მაღალი კონცენტრაცია მოცულობის მარცხენა ნაწილში, C:, - ნაწილაკების დაბალი კონცენტრაცია მარჯვნივ. ნაწილები, ვარდისფერი ზედაპირიარის კონცენტრაციის გრადიენტი დიფუზიურ შრეში. დიფუზიური ნაკადი dm/dt – იხ განტოლება (1)
ნივთიერებები ასევე კარგად ვრცელდება მემბრანებში; ეს ასევე ეხება შედარებით მცირე პოლარულ მოლეკულებს, როგორიცაა ეთანოლი და შარდოვანა, ხოლო შაქარი გადის ლიპიდურ შრეში გაჭირვებით. ამავდროულად, ლიპიდური ფენები პრაქტიკულად გაუვალია დამუხტული მოლეკულებისთვის, მათ შორის არაორგანული იონებისთვისაც კი. არაელექტროლიტებისთვის, დიფუზიის განტოლება (1) ჩვეულებრივ გარდაიქმნება მემბრანის და დიფუზიური ნივთიერების მახასიათებლების ერთში გაერთიანებით. პარამეტრი-გამტარიანობა (P):
dm/dt=P Aდ გ.(3)
ნახ. 1.4 შედარებით ლიპიდური მემბრანის გამტარიანობა (P) სხვადასხვა მოლეკულებისთვის.
დიფუზია მემბრანის ფორებში . პლაზმური მემბრანა (და სხვა უჯრედული მემბრანა) გამტარია არა მხოლოდ ლიპიდური შრის მეშვეობით დიფუზური ნივთიერებებისთვის, არამედ მრავალი იონის, შაქრის, ამინომჟავებისა და ნუკლეოტიდების მიმართ. ეს ნივთიერებები კვეთს მემბრანას წარმოქმნილი ფორებით სატრანსპორტო ცილები,მემბრანაში ჩადგმული. ასეთი ცილების შიგნით არის 1 ნმ-ზე ნაკლები დიამეტრის წყლით სავსე არხი, რომლის მეშვეობითაც მცირე მოლეკულები შეიძლება გავრცელდეს. ისინი მოძრაობენ კონცენტრაციის გრადიენტის გასწვრივ და თუ ისინი ატარებენ მუხტს, მაშინ მათი მოძრაობა არხებში ასევე რეგულირდება მემბრანის პოტენციალით. მემბრანული არხები შედარებით შერჩევითია

ბრინჯი. 1.4.ხელოვნური ლიპიდური ორფენიანი გამტარიანობა სხვადასხვა ნივთიერებისთვის
მოლეკულების ტიპთან მიმართებაში, რომელსაც შეუძლია მათში გავლა, არსებობს, მაგალითად, კალიუმის, ნატრიუმის და კალციუმის არხები, რომელთაგან თითოეული გაუმტარია თითქმის ნებისმიერი იონის მიმართ, გარდა კონკრეტულისა. ასეთი შერჩევითობაარხის კედლებში დამაკავშირებელი ადგილების დატენვის ან სტრუქტურის გამო, რაც ხელს უწყობს კონკრეტული მოლეკულის ტრანსპორტირებას და ხელს უშლის სხვა ნივთიერებების შეღწევას არხში (ნახ. 1.5, ა) .
ქცევის უკან მემბრანული იონური არხებიმისი დაკვირვება მარტივია, რადგან იონების გადაადგილების დროს წარმოქმნილი დენი შეიძლება გაიზომოს და თუნდაც ერთი არხისთვის. ნაჩვენებია, რომ არხები სპონტანურად და მაღალი სიხშირით ცვლის თავის მდგომარეობას ღიადან დახურულში. კალიუმის არხს ახასიათებს დენის პულსები დაახლოებით 2 pA (2 10 -12 A) ამპლიტუდით და რამდენიმე მილიწამის ხანგრძლივობით (იხ. სურ. 2.12, გვ. 37) [3]. ამ პერიოდის განმავლობაში მასში ათობით ათასი იონი გადის. ცილების გადასვლა ერთი კონფორმაციიდან მეორეზე შესწავლილია რენტგენის დიფრაქციით, მოსბაუერის სპექტროსკოპიით და ბირთვული მაგნიტური რეზონანსით (NMR). ამგვარად, პროტეინები ძალიან დინამიური, მოძრავი სტრუქტურებია და პროტეინის გავლით არხი არ არის მხოლოდ ხისტი, წყლით სავსე მილი (სურათი 1.5A), არამედ სწრაფად მოძრავი მოლეკულური ჯგუფებისა და მუხტების ლაბირინთი. ეს არხის დინამიური პასუხი აისახება არხის ენერგეტიკული პროფილი,ნაჩვენებია ნახ. 1.5, B. აქ აბსციზა გვიჩვენებს არხის სიგრძეს გარე ხსნარიდან C 0 იონის კონცენტრაციით და 0 პოტენციალით შიდა ხსნარამდე C 1 კონცენტრაციით და E პოტენციალით. y ღერძი.

ბრინჯი. 1.5.ა. ცილის სქემა, რომელიც ქმნის კალიუმის არხს, რომელიც ჩაშენებულია პლაზმური მემბრანის ლიპიდურ ორ ფენაში. არხის „კედელზე“ ოთხი უარყოფითი მუხტი ფიქსირდება. ბ. არხის სქემატური ენერგეტიკული პროფილი, რომელიც ნაჩვენებია ნახ. ა. y ღერძზე გამოსახულია არხის გასავლელად საჭირო კინეტიკური ენერგიის მნიშვნელობები; აბსცისის ღერძის გასწვრივ, გარსის შიდა და გარე ზედაპირებს შორის დაშორებები. ენერგეტიკული მინიმუმები შეესაბამება არხის კედელში დადებითად დამუხტული იონების შეკავშირების ადგილებს ფიქსირებული უარყოფითი მუხტით. ენერგიის მაქსიმუმი შეესაბამება არხში დიფუზიის დაბრკოლებებს. ვარაუდობენ, რომ არხის ცილის კონფორმაცია სპონტანურად ირხევა; ენერგეტიკული პროფილების ვარიანტები ნაჩვენებია მყარი და წყვეტილი ხაზებით; ეს რხევები მნიშვნელოვნად უწყობს ხელს იონების შეკავშირებას ენერგეტიკული ბარიერის გადალახვისას (მაგრამ ცვლილებებით)
ნაჩვენებია იონის ენერგეტიკული დონეები არხის დამაკავშირებელ ადგილებში; დიაგრამაზე პიკი წარმოადგენს გამტარიანობის ბარიერს, რომელიც უნდა გადალახოს იონურმა ენერგიამ არხში შეღწევის მიზნით, ხოლო გრაფიკის „დაწევა“ წარმოადგენს შედარებით სტაბილურ მდგომარეობას (დაკავშირებას). ენერგიის პიკის დაბრკოლების მიუხედავად, იონს შეუძლია შეაღწიოს არხში, თუ ენერგეტიკული პროფილი სპონტანურად ციკლდება; ამრიგად, იონი შეიძლება მოულოდნელად გამოჩნდეს ენერგიის პიკის "მეორე მხარეს" და გააგრძელოს გადაადგილება უჯრედში. იონის მუხტის, ზომისა და ჰიდრატაციის ხარისხზე და არხის კედლების სტრუქტურებთან შეკავშირების შესაძლებლობიდან გამომდინარე, არხის ენერგეტიკული პროფილი განსხვავდება სხვადასხვა იონისთვის, რამაც შეიძლება აიხსნას არხების ცალკეული ტიპის სელექციურობა.
იონების დიფუზიური წონასწორობა . მემბრანული არხებით სხვადასხვა იონების დიფუზიამ უნდა გამოიწვიოს კონცენტრაციების განსხვავებების აღმოფხვრა ექსტრა და უჯრედშიდა გარემოს შორის. თუმცა, როგორც ჩანს ცხრილიდან. 1.1, ასეთი განსხვავებები შენარჩუნებულია, ამიტომ გარკვეული უნდა იყოს წონასწორობადიფუზიასა და მემბრანის გასწვრივ სხვა სატრანსპორტო პროცესებს შორის. შემდეგი ორი ნაწილი განიხილავს გზებს, რომლითაც ასეთი წონასწორობა დამყარებულია. იონების შემთხვევაში დიფუზიის წონასწორობაზე გავლენას ახდენს მათი მუხტი. დაუმუხტი მოლეკულების დიფუზია უზრუნველყოფილია კონცენტრაციის სხვაობითდკ და როდესაც კონცენტრაციები გათანაბრდება, ფაქტობრივი ტრანსპორტი ჩერდება. დამუხტულ ნაწილაკებზე დამატებით გავლენას ახდენს ელექტრული ველი. მაგალითად, როდესაც კალიუმის იონი გამოდის უჯრედიდან მისი კონცენტრაციის გრადიენტის გასწვრივ, ის ატარებს ერთ დადებით მუხტს. ამრიგად, უჯრედშიდა გარემო უფრო უარყოფითად დამუხტული ხდება, რაც იწვევს პოტენციურ განსხვავებას მემბრანაში. უჯრედშიდა უარყოფითი მუხტი ხელს უშლის ახალი კალიუმის იონების უჯრედიდან გასვლას და ის იონები, რომლებიც მაინც ტოვებენ უჯრედს, კიდევ უფრო გაზრდის მუხტს მემბრანაზე. კალიუმის იონების ნაკადი ჩერდება, როდესაც ელექტრული ველის მოქმედება ანაზღაურებს დიფუზიურ წნევას კონცენტრაციების სხვაობის გამო. იონები აგრძელებენ მემბრანის გავლას, მაგრამ თანაბარი რაოდენობით ორივე მიმართულებით. მაშასადამე, მემბრანაზე იონების კონცენტრაციებში მოცემული სხვაობისთვის არსებობს წონასწორობის პოტენციალიეიონი რომლის დროსაც მემბრანაში იონების დინება ჩერდება. წონასწორობის პოტენციალი ადვილად შეიძლება განისაზღვროს გამოყენებით ნერნსტის განტოლებები:
ეიონი= რთ/ ზფ* ლნგარეთ/ C in(4)
სადაც რ არის აირის მუდმივი, T არის აბსოლუტური ტემპერატურა, z არის იონის ვალენტობა (უარყოფითი ანიონებისთვის)გარეთ არის უჯრედგარე იონის კონცენტრაცია, C in არის იონის უჯრედშიდა კონცენტრაცია,ფ ფარადეის ნომერი. თუ განტოლებაში ჩავანაცვლებთ მუდმივებს, მაშინ სხეულის ტემპერატურაზე (T = 310 K) კალიუმის იონების წონასწორობის პოტენციალი E K უდრის:
ეკ= –61 მB ჟურნალი /(5)
თუ [ K + out ]/[ K + in ] = 39, როგორც ქვემოთ მოცემულია ცხრილიდან. 1.1, მაშინ
Ek= -61 m B log 39= -97 mV.
მართლაც, აღმოჩნდა, რომ ყველა უჯრედს აქვს მემბრანის პოტენციალი;ძუძუმწოვრების კუნთოვან უჯრედებში მისი დონე დაახლოებით -90 მვ-ია. იონების პირობებისა და შედარებითი კონცენტრაციიდან გამომდინარე, უჯრედებს შეიძლება ჰქონდეთ მემბრანული პოტენციალი -40-დან -120 მვ-მდე. ზემოთ მოყვანილი მაგალითის უჯრედისთვის (ცხრილი 1.1) დასვენების პოტენციალი,უდრის დაახლოებით -90 მვ-ს, მიუთითებს იმაზე, რომ კალიუმის იონების ნაკადები მემბრანულ არხებში დაახლოებით წონასწორობაშია. ეს გასაკვირი არ არის, რადგან მოსვენებულ მემბრანაში კალიუმის არხების ღია მდგომარეობა ყველაზე სავარაუდოა; მემბრანა ყველაზე გამტარია კალიუმის იონების მიმართ. თუმცა, მემბრანის პოტენციალი განისაზღვრება სხვა იონების ნაკადითაც.
სიმარტივე, რომლითაც დაუმუხტავ ნაწილაკებს შეუძლიათ მემბრანაში დიფუზია, რაოდენობრივია (3) განტოლებაში. დამუხტული ნაწილაკების გამტარიანობა აღწერილია ოდნავ უფრო რთული განტოლებით:
პ= მ რთ/ dF(6)
სადაც მარის მემბრანაში იონის მობილურობა,დ - მემბრანის სისქე, a R, T და F ცნობილი თერმოდინამიკური მუდმივები. ამ გზით განსაზღვრული სხვადასხვა იონების გამტარიანობის მნიშვნელობები შეიძლება გამოყენებულ იქნას მემბრანის პოტენციალის გამოსათვლელადემ როდესაც კალიუმის, ნატრიუმის და ქლორიდის იონები გადიან მემბრანაში ერთდროულად (გამტარიანობით P K, P Na და P Cl შესაბამისად). ვარაუდობენ, რომ პოტენციალი მემბრანაში ერთნაირად ეცემა, ისე რომ ველის სიძლიერე მუდმივია. ამ შემთხვევაში, ეს ეხება გოლდმანის განტოლება, ან მუდმივი ველის განტოლება :
ეm= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
უჯრედის მემბრანების უმეტესობისთვის Pკ დაახლოებით 30-ჯერ აღემატება რნა (იხილეთ აგრეთვე ნაწილი 1.3). შედარებითი ღირებულება PCl ძალიან განსხვავდება; მრავალი მემბრანისთვის PCl მცირეა რ-თან შედარებითკ თუმცა სხვებისთვის (მაგ. ჩონჩხის კუნთებში) PCl , ბევრად აღემატება რკ.
აქტიური ტრანსპორტი, ნატრიუმის ტუმბო . წინა განყოფილება აღწერს იონების პასიურ დიფუზიას და მემბრანულ პოტენციალს მოცემული უჯრედშიდა და უჯრედგარე იონების კონცენტრაციებში. თუმცა, ამ პროცესის შედეგად, უჯრედის შიგნით იონების კონცენტრაცია ავტომატურად არ სტაბილიზდება, რადგან მემბრანა
პოტენციალი ოდნავ უფრო ელექტროუარყოფითია ვიდრე E K, და ბევრად მეტი ენა (დაახლოებით +60 მვ). დიფუზიის გამო, იონების, სულ მცირე, კალიუმის და ნატრიუმის უჯრედშიდა კონცენტრაციები უნდა გათანაბრდეს უჯრედგარეთ. იონური გრადიენტის სტაბილურობა მიიღწევა აქტიური ტრანსპორტით: მემბრანის ცილები იონებს მემბრანის გასწვრივ ატარებენ ელექტრული და (ან) კონცენტრაციის გრადიენტების წინააღმდეგ, ამისთვის მოიხმარენ მეტაბოლურ ენერგიას. აქტიური ტრანსპორტის ყველაზე მნიშვნელოვანი პროცესი სამუშაოანა/კ - ტუმბო, რომელიც არსებობს თითქმის ყველა უჯრედში;
ტუმბო ტუმბოს ნატრიუმის იონებს უჯრედიდან, ხოლო ერთდროულად ტუმბოს კალიუმის იონებს უჯრედში. ეს უზრუნველყოფს ნატრიუმის იონების და მაღალი კალიუმის დაბალ უჯრედშიდა კონცენტრაციას (ცხრილი 1.1). მემბრანაზე ნატრიუმის იონების კონცენტრაციის გრადიენტს აქვს სპეციფიკური ფუნქციები, რომლებიც დაკავშირებულია ინფორმაციის გადაცემასთან ელექტრული იმპულსების სახით (იხ. პუნქტი 2.2), აგრეთვე სხვა აქტიური სატრანსპორტო მექანიზმების შენარჩუნებასთან და უჯრედის მოცულობის რეგულირებასთან (იხ. ქვემოთ). ამიტომ გასაკვირი არ არის, რომ უჯრედის მიერ მოხმარებული ენერგიის 1/3-ზე მეტი იხარჯება Na/K ტუმბოზე, ხოლო ზოგიერთ ყველაზე აქტიურ უჯრედში ენერგიის 70%-მდე იხარჯება მის მუშაობაზე.
Na/K სატრანსპორტო ცილა არის ATPase. მემბრანის შიდა ზედაპირზე ის არღვევს ATP-ს ADP-ად და ფოსფატად (ნახ. 1.6). ერთი ATP მოლეკულის ენერგია გამოიყენება უჯრედიდან სამი ნატრიუმის იონის გადასატანად და ერთდროულად ორი კალიუმის იონის უჯრედში, ანუ მთლიანობაში ერთი ციკლიდან ერთი დადებითი მუხტი იხსნება უჯრედიდან. ამრიგად Na/K ტუმბო არის ელექტროგენური(ქმნის ელექტრულ დენს მემბრანის მეშვეობით), რაც იწვევს მემბრანის პოტენციალის ელექტრონეგატიურობის ზრდას დაახლოებით 10 მვ-ით. სატრანსპორტო ცილა ასრულებს ამ ოპერაციას მაღალი სიჩქარით: 150-დან 600-მდე ნატრიუმის იონამდე წამში. ცნობილია სატრანსპორტო ცილის ამინომჟავების თანმიმდევრობა, მაგრამ ამ რთული გაცვლის ტრანსპორტირების მექანიზმი ჯერ არ არის ნათელი. ეს პროცესი აღწერილია ცილებით ნატრიუმის ან კალიუმის იონების გადაცემის ენერგეტიკული პროფილების გამოყენებით (ნახ. 1.5.5). ამ პროფილების ცვლილების ბუნებით, რომელიც დაკავშირებულია სატრანსპორტო ცილის კონფორმაციის მუდმივ ცვლილებებთან (პროცესი, რომელიც მოითხოვს ენერგიას), შეიძლება ვიმსჯელოთ გაცვლის სტოქიომეტრიაზე: კალიუმის ორი იონი იცვლება სამ ნატრიუმის იონზე.
Na/K-ტუმბო, იზოლირებულივით Na+ /K +-დამოკიდებული მემბრანული ატფ-აზა, სპეციალურად ინჰიბირებული საგულე გლიკოზიდი უაბაინი (სტროფანტინი). ვინაიდან Na/K ტუმბოს მუშაობა მრავალსაფეხურიანი ქიმიური რეაქციაა, ის, ისევე როგორც ყველა ქიმიური რეაქცია, დიდად არის დამოკიდებული ტემპერატურაზე, რომელიც

ბრინჯი. 1.6.Na/K-ტუმბო-ATP-აზას სქემა (ჩაძირულია პლაზმური მემბრანის ლიპიდურ ორ შრეში), რომელიც ერთ ციკლში უჯრედიდან სამ Na + იონს გამოაქვს პოტენციური და კონცენტრაციის გრადიენტების საწინააღმდეგოდ და უჯრედში შემოაქვს ორი K იონი. + . ამ პროცესის დროს, ერთი ATP მოლეკულა იყოფა ADP და ფოსფატად. დიაგრამაზე ATPase ნაჩვენებია დიმერის სახით, რომელიც შედგება დიდი (ფუნქციური) და მცირე ქვედანაყოფისგან; მემბრანაში ის არსებობს როგორც ტეტრამერი, რომელიც წარმოიქმნება ორი დიდი და ორი პატარა ქვედანაყოფით
ნაჩვენებია ნახ. 1.7. აქ ნატრიუმის იონების ნაკადი კუნთოვანი უჯრედებიდან ნაჩვენებია დროსთან მიმართებაში; ეს პრაქტიკულად ექვივალენტურია ნატრიუმის იონების ნაკადის შუამავლობით Na/K ტუმბოს მუშაობით, რადგან ნატრიუმის იონების პასიური ნაკადი კონცენტრაციისა და პოტენციური გრადიენტების მიმართ ძალიან მცირეა. თუ პრეპარატი გაცივდა დაახლოებით 18°C-ით, მაშინ ნატრიუმის იონების ნაკადი უჯრედიდან სწრაფად შემცირდება 15-ჯერ და გაცხელებისთანავე იგი აღდგება პირვანდელ დონეზე. უჯრედიდან ნატრიუმის იონების ნაკადის ასეთი შემცირება რამდენჯერმე აღემატება იმას, რაც შეესაბამება დიფუზიის პროცესის ტემპერატურულ დამოკიდებულებას ან მარტივ ქიმიურ რეაქციას. მსგავსი ეფექტი შეინიშნება დინიტროფენოლის (DNP) მოწამვლის შედეგად მეტაბოლური ენერგიის დაქვეითებისას (სურ. 1.7.5). ამიტომ ნატრიუმის იონების ნაკადს უჯრედიდან უზრუნველყოფს ენერგიაზე დამოკიდებული რეაქცია – აქტიური ტუმბო. ტუმბოს კიდევ ერთი მახასიათებელი, ტემპერატურისა და ენერგიის მნიშვნელოვან დამოკიდებულებასთან ერთად, არის გაჯერების დონის არსებობა (როგორც ყველა სხვა ქიმიური რეაქცია); ეს ნიშნავს, რომ ტუმბოს სიჩქარე განუსაზღვრელი ვადით არ შეიძლება გაიზარდოს ტრანსპორტირებული იონების კონცენტრაციის მატებასთან ერთად (ნახ. 1.8). ამის საპირისპიროდ, პასიურად დიფუზიური ნივთიერების ნაკადი იზრდება კონცენტრაციების სხვაობის პროპორციულად დიფუზიის კანონის შესაბამისად (განტოლებები 1 და 2).

ბრინჯი. 1.7. A, B.აქტიური ტრანსპორტია + . Y-ღერძი:რადიოაქტიური 24 Na + გადინება უჯრედიდან (იმპ./წთ). აბსცისის ღერძი:დრო ექსპერიმენტის დაწყებიდან. მაგრამ.უჯრედი გაცივებულია 18,3°C-დან 0,5°C-მდე; ნაკადი Na+ ამ პერიოდში უჯრედიდან გასვლა იზღუდება. ბ.უჯრედიდან Na + ნაკადის ჩახშობა დინიტროფენოლით (DNF) 0,2 მმოლ/ლ კონცენტრაციით (შეცვლილი)
Na/K ტუმბოს გარდა, პლაზმური მემბრანა შეიცავს მინიმუმ კიდევ ერთ ტუმბოს - კალციუმი;ეს ტუმბო ტუმბოს კალციუმის იონებს (Ca 2+) უჯრედიდან და მონაწილეობს მათი უჯრედშიდა კონცენტრაციის უკიდურესად დაბალ დონეზე შენარჩუნებაში (ცხრილი 1.1). კალციუმის ტუმბო ძალიან მაღალი სიმკვრივით იმყოფება კუნთოვანი უჯრედების სარკოპლაზმურ რეტიკულუმში, რომლებიც აგროვებენ კალციუმის იონებს ATP მოლეკულების დაშლის შედეგად (იხ. თავი 4).
Na/K ტუმბოს ეფექტი მემბრანის პოტენციალსა და უჯრედის მოცულობაზე . ნახ. 1.9 გვიჩვენებს მემბრანის დენის სხვადასხვა კომპონენტს და გვიჩვენებს იონების უჯრედშიდა კონცენტრაციებს, რომლებიც

ბრინჯი. 1.8.თანაფარდობა მოლეკულების ტრანსპორტირების სიჩქარესა და მათ კონცენტრაციას შორის (არხში შესვლის ან ტუმბოს შებოჭვის ადგილზე) არხში დიფუზიის დროს ან სატუმბი ტრანსპორტირების დროს. ეს უკანასკნელი გაჯერებულია მაღალი კონცენტრაციით (მაქსიმალური მაჩვენებელი, Vmax ) მნიშვნელობა აბსციზაზე, რომელიც შეესაბამება ტუმბოს მაქსიმალური სიჩქარის ნახევარს ( Vmax /2), არის წონასწორული კონცენტრაცია TO მ

ბრინჯი. 1.9.დიაგრამა, რომელიც აჩვენებს Na+ კონცენტრაციას , K+ და Cl- უჯრედის შიგნით და გარეთ და ამ იონების უჯრედის მემბრანაში შეღწევის გზები (სპეციფიკური იონური არხებით ან Na/K ტუმბოს დახმარებით. მოცემული კონცენტრაციის გრადიენტებით, წონასწორობის პოტენციალი E Na, E K და E C l - უტოლდება მითითებულებს, მემბრანულ პოტენციალსემ = – 90 მვ
უზრუნველყოს მათი არსებობა. კალიუმის იონების გამავალი დენი შეინიშნება კალიუმის არხებით, რადგან მემბრანის პოტენციალი გარკვეულწილად უფრო ელექტროპოზიტიურია, ვიდრე კალიუმის იონების წონასწორობის პოტენციალი. ნატრიუმის არხების საერთო გამტარობა გაცილებით დაბალია, ვიდრე კალიუმის არხები; ნატრიუმის არხები იხსნება ბევრად უფრო იშვიათად, ვიდრე კალიუმის არხები მოსვენების დროს; თუმცა, ნატრიუმის იონების დაახლოებით იგივე რაოდენობა შედის უჯრედში, რამდენიც კალიუმის იონებს ტოვებენ, რადგან უჯრედში ნატრიუმის იონების დიფუზიისთვის საჭიროა დიდი კონცენტრაცია და პოტენციური გრადიენტები. Na/K ტუმბო უზრუნველყოფს პასიური დიფუზიის დენების იდეალურ კომპენსაციას, რადგან ის გადააქვს ნატრიუმის იონებს უჯრედიდან და კალიუმის იონებს მასში. ამრიგად, ტუმბო ელექტროგენულია უჯრედში და მის გარეთ გადატანილი მუხტების რაოდენობის განსხვავების გამო, რაც მისი მუშაობის ნორმალური სიჩქარით ქმნის მემბრანულ პოტენციალს დაახლოებით 10. mV უფრო ელექტროუარყოფითია, ვიდრე მხოლოდ პასიური იონის დენებისაგან რომ წარმოქმნილიყო (იხ. განტოლება 7). შედეგად, მემბრანის პოტენციალი უახლოვდება კალიუმის წონასწორობის პოტენციალს, რაც ამცირებს კალიუმის იონების გაჟონვას. Na აქტიურობა/K-ტუმბო რეგულირდება ნატრიუმის იონების უჯრედშიდა კონცენტრაციით. ტუმბოს სიჩქარე ნელდება უჯრედიდან გამოსატანი ნატრიუმის იონების კონცენტრაციის დაქვეითებისას (ნახ. 1.8), ასე რომ, ტუმბოს მუშაობა და ნატრიუმის იონების ნაკადი უჯრედში ერთმანეთს აბალანსებს და ინარჩუნებს უჯრედშიდა კონცენტრაციას. ნატრიუმის იონები დაახლოებით 10 მმოლ/ლ დონეზე.
ტუმბოსა და მემბრანის პასიურ დენებს შორის ბალანსის შესანარჩუნებლად საჭიროა მეტი Na/K-ტუმბოს მოლეკულა, ვიდრე კალიუმის და ნატრიუმის იონების არხის ცილები. როდესაც არხი ღიაა, მასში ათიათასობით იონი გადის რამდენიმე მილიწამში (იხ. ზემოთ), და რადგან არხი ჩვეულებრივ იხსნება წამში რამდენჯერმე, ამ დროის განმავლობაში მასში სულ 10 5 იონზე მეტი გადის. ერთი ტუმბოს ცილა წამში რამდენიმე ასეულ ნატრიუმის იონს მოძრაობს, ამიტომ პლაზმური მემბრანა უნდა შეიცავდეს დაახლოებით 1000-ჯერ მეტ ტუმბოს მოლეკულას, ვიდრე არხის მოლეკულებს. არხის დენების გაზომვამ აჩვენა საშუალოდ ერთი კალიუმის და ერთი ნატრიუმის ღია არხი 1 μm 2 მემბრანაზე; აქედან გამომდინარეობს, რომ დაახლოებით 1000 Na/K ტუმბოს მოლეკულა უნდა იყოს იმავე სივრცეში, ე.ი. მათ შორის მანძილი საშუალოდ 34 ნმ; სატუმბი ცილის, როგორც არხის პროტეინის დიამეტრი არის 8-10 ნმ. ამრიგად, მემბრანა საკმარისად მჭიდროდ არის გაჯერებული სატუმბი მოლეკულებით
ის ფაქტი, რომ უჯრედში ნატრიუმის იონების გადინება და უჯრედიდან კალიუმის იონები კომპენსირდება ტუმბოს მუშაობით, აქვს კიდევ ერთი შედეგი, რომელიც მოიცავს სტაბილური ოსმოსური წნევის შენარჩუნებას და მუდმივი მოცულობა.უჯრედის შიგნით არის დიდი ანიონების მაღალი კონცენტრაცია, ძირითადად ცილები (A - ცხრილი 1.1), რომლებსაც არ შეუძლიათ მემბრანაში შეღწევა (ან ძალიან ნელა შეაღწიონ მასში) და, შესაბამისად, უჯრედის შიგნით ფიქსირებულ კომპონენტს წარმოადგენენ. ამ ანიონების მუხტის დასაბალანსებლად საჭიროა კათიონის თანაბარი რაოდენობა. Na/K ტუმბოს მოქმედების გამო ეს კათიონები ძირითადად კალიუმის იონებია. იონების უჯრედშიდა კონცენტრაციის მნიშვნელოვანი ზრდა შეიძლება მოხდეს მხოლოდ ანიონების კონცენტრაციის ზრდით C1-ის ნაკადის გამო - კონცენტრაციის გრადიენტის გასწვრივ უჯრედში (ცხრილი 1.1), მაგრამ მემბრანის პოტენციალი ეწინააღმდეგება ამას. შემომავალი დენი Cl- შეინიშნება მხოლოდ ქლორიდის იონების წონასწორობის პოტენციალის მიღწევამდე; ეს შეინიშნება, როდესაც ქლორიდის იონის გრადიენტი თითქმის საპირისპიროა კალიუმის იონის გრადიენტთან, ვინაიდან ქლორიდის იონები უარყოფითად არის დამუხტული (განტოლება 4). ამრიგად, იქმნება ქლორიდის იონების დაბალი უჯრედშიდა კონცენტრაცია, რაც შეესაბამება კალიუმის იონების დაბალ უჯრედგარე კონცენტრაციას. შედეგი არის უჯრედში იონების საერთო რაოდენობის შეზღუდვა. თუ მემბრანის პოტენციალი ეცემა Na/K ტუმბოს დაბლოკვისას, მაგალითად, ანოქსიის დროს, მაშინ ქლორიდის იონების წონასწორობის პოტენციალი მცირდება და შესაბამისად იზრდება ქლორიდის იონების უჯრედშიდა კონცენტრაცია. მუხტების ბალანსის აღდგენით, კალიუმის იონებიც შედიან უჯრედში; უჯრედში იზრდება იონების საერთო კონცენტრაცია, რაც ზრდის ოსმოსურ წნევას; ეს აიძულებს წყალს შევიდეს უჯრედში. უჯრედი ადიდებს. ეს შეშუპება ჩანს in vivo ენერგიის ნაკლებობის პირობებში.
კონცენტრაციის გრადიენტი ნა + როგორც მემბრანული ტრანსპორტირების მამოძრავებელი ძალა . Na/K–ტუმბოს მნიშვნელობა უჯრედისთვის არ შემოიფარგლება მემბრანის გასწვრივ ნორმალური K+ და Na+ გრადიენტების სტაბილიზირებით. მემბრანის გრადიენტში შენახული ენერგია Na+ , ხშირად გამოიყენება სხვა ნივთიერებების მემბრანული ტრანსპორტირების უზრუნველსაყოფად. მაგალითად, ნახ. 1.10 აჩვენებს "სიმპორტს" Na+ და შაქრის მოლეკულები უჯრედში. მემბრანის სატრანსპორტო ცილა ატარებს შაქრის მოლეკულას უჯრედში კონცენტრაციის გრადიენტის წინააღმდეგაც კი, ხოლო Na + მოძრაობს კონცენტრაციისა და პოტენციური გრადიენტის გასწვრივ, რაც უზრუნველყოფს ენერგიას. შაქრის ტრანსპორტირება.შაქრის ასეთი ტრანსპორტი მთლიანად დამოკიდებულია მაღალი გრადიენტის არსებობაზე Na+ ; თუ უჯრედშიდა კონცენტრაცია Na+ მნიშვნელოვნად იზრდება, შემდეგ შაქრის ტრანსპორტირება ჩერდება. სხვადასხვასთვისგ ახაროვი, არსებობს სხვადასხვა სიმპორტის სისტემა. ამინომჟავების ტრანსპორტირებატრანსპორტის მსგავს გალიაშიგ ახაროვი ნაჩვენებია ნახ. 1.10; იგი ასევე უზრუნველყოფილია გრადიენტით Na+ , არსებობს სულ მცირე ხუთი განსხვავებული სიმპორტის სისტემა, თითოეული სპეციალიზირებულია დაკავშირებული ამინომჟავების ერთი ჯგუფისთვის.
სიმპორტის სისტემების გარდა, არსებობს ასევე "ანტიპორტი".ერთ-ერთი მათგანი, მაგალითად, ერთი ციკლის განმავლობაში გადააქვს ერთი კალციუმის იონი უჯრედიდან სამი შემომავალი ნატრიუმის იონების სანაცვლოდ (ნახ. 1.10). ენერგია ამისთვის ტრანსპორტი Ca 2+წარმოიქმნება კონცენტრაციისა და პოტენციური გრადიენტის გასწვრივ სამი ნატრიუმის იონების შეყვანის გამო. ეს ენერგია საკმარისია (დასვენების პოტენციალის დროს) მაღალი კალციუმის იონური გრადიენტის შესანარჩუნებლად (უჯრედის შიგნით 10-7 მოლ/ლ-ზე ნაკლები, უჯრედის გარეთ დაახლოებით 2 მმოლ/ლ-მდე).
ენდო- და ეგზოციტოზი . გარკვეული ნივთიერებებისთვის, რომლებიც შედის უჯრედში ან უნდა მოიხსნას

ბრინჯი. 1.10.მემბრანის ლიპიდურ ორ ფენაში ჩადებული პროტეინები შუამავლობენ გლუკოზის და Na + უჯრედში იმპორტს, აგრეთვე Ca2+/Na+ -ანტიპორტი, რომელშიც მამოძრავებელი ძალა არის Na + გრადიენტი უჯრედის მემბრანაზე
მისგან არ არის სატრანსპორტო არხები; ასეთი ნივთიერებები მოიცავს, მაგალითად, ცილებს და ქოლესტერინს. მათ შეუძლიათ პლაზმური მემბრანის გავლით ვეზიკულებიან ბუშტები,ენდო- და ეგზოციტოზით. ნახ. 1.11 გვიჩვენებს ამ პროცესების ძირითად მექანიზმებს. ეგზოციტოზის დროს, გარკვეული ორგანელები (იხ. ქვემოთ) ქმნიან ბუშტუკებს, რომლებიც სავსეა უჯრედიდან ამოღებული ნივთიერებით, როგორიცაა ჰორმონები ან უჯრედგარე ფერმენტები. როდესაც ასეთი ვეზიკულები აღწევს პლაზმურ მემბრანას, მათი ლიპიდური მემბრანა ერწყმის მას, რითაც საშუალებას აძლევს შიგთავსს გაექცეს გარე გარემოში. საპირისპირო პროცესში, ენდოციტოზის დროს, პლაზმური მემბრანა ინვაგინარდება, წარმოქმნის ორმოს, რომელიც შემდეგ ღრმავდება და იხურება, წარმოიქმნება უჯრედშორისი სითხით და ზოგიერთი მაკრომოლეკულით სავსე უჯრედშიდა ვეზიკულა. ამ მემბრანის შერწყმისა და ვეზიკულის დახურვის უზრუნველსაყოფად, ციტოჩონჩხის კონტრაქტული ელემენტები მოქმედებენ თავად მემბრანებთან ერთად (იხ. ქვემოთ). ენდოციტოზი ყოველთვის არ გულისხმობს უჯრედშიდა უჯრედული საშუალების უბრალოდ შეყვანას. უჯრედის მემბრანა შეიცავს, ხშირად სპეციალიზებულ ჯგუფებად ორგანიზებულ, სპეციფიკურ რეცეპტორებს მაკრომოლეკულებისთვის, როგორიცაა ინსულინი ან ანტიგენები. მას შემდეგ, რაც ეს მაკრომოლეკულები დაუკავშირდებიან მათ რეცეპტორებს, ენდოციტოზი ხდება რეცეპტორის მიმდებარე მემბრანის რეგიონში და მაკრომოლეკულა შერჩევით ტრანსპორტირდება უჯრედში (ნახ. 1.12, B).
ენდო- და ეგზოციტოზი მუდმივად ხდება უჯრედებში. მემბრანული მასალის მოცირკულირე რაოდენობა მნიშვნელოვანია; 1 საათის განმავლობაში მაკროფაგი ბუშტუკების სახით შთანთქავს მისი ციტოპლაზმური მემბრანის ზედაპირის ორჯერ მეტს. უჯრედების უმეტესობაში მემბრანული მასალის ბრუნვა არც თუ ისე ინტენსიურია, მაგრამ მაინც მნიშვნელოვანი უნდა იყოს.

ბრინჯი. 1.11.ეგზოციტოზი და ენდოციტოზი. ზემოთ:უჯრედშიდა ვეზიკულა ერწყმის პლაზმური მემბრანის ლიპიდურ ორ ფენას და იხსნება უჯრედგარე სივრცეში. ამ პროცესს ეგზოციტოზი ეწოდება. ბოლოში:პლაზმური მემბრანა შემოიჭრება მცირე ფართობზე და აშორებს უჯრედგარე მასალით სავსე ვეზიკულას. ამ პროცესს ენდოციტოზს უწოდებენ.
1.3. ნივთიერებების ტრანსპორტირება უჯრედში
ენდო- და ეგზოციტოზი არის არა მხოლოდ უჯრედის მემბრანის მეშვეობით ნივთიერებების ტრანსპორტირების პროცესები, არამედ მემბრანული გაცვლის პროცესები - თავად უჯრედის სტრუქტურული კომპონენტები. ამ ნაწილში განხილვის საგანია სხვა მსგავსი სატრანსპორტო პროცესები უჯრედსა და მის ორგანელებში.

ბრინჯი. 1.12. A-B.პროცესების სქემა, მათ შორის ეგზო-და ენდოციტოზი. მაგრამ.მარცვლოვან ენდოპლაზმურ რეტიკულუმში სინთეზირებული ცილა გოლჯის აპარატის მეშვეობით პლაზმურ მემბრანაში ტრანსპორტირდება, სადაც ის გამოიყოფა ეგზოციტოზის გზით. ბ. LDL (დაბალი სიმკვრივის ლიპოპროტეინების) ნაწილაკებთან შეკრული ქოლესტერინი მიმაგრებულია პლაზმურ მემბრანაზე, იწვევს მემბრანის ამ რეგიონში ენდოციტური ვეზიკულის წარმოქმნას და ტრანსპორტირდება ლიზოსომებში, სადაც ის გამოიყოფა. IN.ენდოციტოზის დროს დაჭერილი უჯრედგარე მასალა (სურათზე მარჯვნივ),ტრანსპორტირება უჯრედში ვეზიკულებში, ან ვეზიკულებში და გამოიყოფა ეგზოციტოზის საშუალებით (სურათზე მარცხენა)
დიფუზია . ბუნებრივია, ციტოზოლში კონცენტრაციის სხვაობა აღმოიფხვრება დიფუზიით; იგივე ეხება ორგანელებში შემავალ სითხეებს. გახსნილი ცილის მაღალი კონცენტრაციის გამო აქ დიფუზია გაცილებით ნელა მიმდინარეობს, ვიდრე წყალში. ლიპიდური მემბრანები - უჯრედის ირგვლივ და ორგანელებში - არის ორგანზომილებიანი სითხეები, რომლებშიც ხდება დიფუზია. ლიპიდები მემბრანულ ორ შრეში დიფუზობენ საკუთარ შრეში, იშვიათად გადადიან ერთიდან მეორეზე. მათში ჩაძირული ცილებიც საკმაოდ მოძრავია; ისინი ბრუნავენ მემბრანის პერპენდიკულარულ ღერძზე ან დიფუზობენ ლატერალურად ძალიან განსხვავებული დიფუზიის მუდმივებით, 2-10000-ჯერ უფრო ნელა, ვიდრე ფოსფოლიპიდები. ასე რომ, თუ ზოგიერთი ცილა თავისუფლად მოძრაობს ლიპიდურ შრეში და იმავე სიჩქარით, როგორც თავად ლიპიდური მოლეკულები, მაშინ სხვები იკვრება, ე.ი. საკმაოდ მტკიცედ არის დაკავშირებული ციტოჩონჩხთან. მემბრანაში არის სპეციფიკური ცილების "მუდმივი" აგრეგატები, როგორიცაა ნერვული უჯრედების პრე- და პოსტსინაფსური სტრუქტურები. თავისუფლად მოძრავი ცილების დემონსტრირება შესაძლებელია ფლუორესცენტურ საღებავებთან მათი შეკავშირებით, რომლებიც ანათებენ მემბრანის მცირე უბნის მოკლე ციმციმებით ხანმოკლე განათებით. ასეთი ექსპერიმენტები აჩვენებს, რომ 1 წუთზე ნაკლებ დროში საღებავთან შეკრული ცილები თანაბრად ნაწილდება მემბრანაზე 10 მკმ-მდე მანძილზე.
აქტიური ტრანსპორტი ორგანულ მემბრანებში .
აქტიური ტრანსპორტის პროცესები, რომლებიც მნიშვნელოვან როლს თამაშობენ პლაზმური მემბრანის ფუნქციონირებაში, ასევე ხდება უჯრედის შიგნით, ორგანელების მემბრანებში. სხვადასხვა ორგანელების სპეციფიკური შინაარსი იქმნება ნაწილობრივ შინაგანი სინთეზით და ნაწილობრივ ციტოზოლიდან აქტიური ტრანსპორტით. ამ უკანასკნელის ერთ-ერთი მაგალითია ზემოთ ნახსენები Ca 2+ ტუმბო კუნთოვანი უჯრედების სარკოპლაზმურ რეტიკულუმში. განსაკუთრებით საინტერესოა, რომ მიტოქონდრიებში ატფ-ის სინთეზის შემთხვევაში, საპირისპირო პრინციპი მოქმედებს პლაზმური მემბრანის ატფ-აზას ტუმბოებში (ნახ. 1.6). ATP სინთეზის დროს ჟანგვითი მეტაბოლიზმი იწვევს ციცაბო გრადიენტის ფორმირებას H+ შიდა გარსებზე. ეს გრადიენტი არის მამოძრავებელი ძალა პროცესისთვის, რომელიც საპირისპიროა აქტიური მოლეკულური ტრანსპორტის სატუმბი ციკლისგან: H + იონები მემბრანის გასწვრივ მოძრაობენ გრადიენტის გასწვრივ და ამის შედეგად გამოთავისუფლებული ენერგია უზრუნველყოფს ATP-ის სინთეზს ADP-დან და ფოსფატიდან. . შედეგად მიღებული ATP, თავის მხრივ, უზრუნველყოფს უჯრედს ენერგიას, მათ შორის აქტიური ტრანსპორტისთვის.
ტრანსპორტირება ვეზიკულებში . უჯრედს აქვს დიდი რაოდენობით ორგანელები და ასოცირებული ვეზიკულები (ნახ. 1.1). ეს ორგანელები და განსაკუთრებით ვეზიკულები მუდმივ მოძრაობაში არიან და გადააქვთ მათი შინაარსი სხვა ორგანელებში ან პლაზმურ მემბრანაში. ბუშტუკებს ასევე შეუძლიათ უჯრედის მემბრანიდან ორგანელებში გადატანა, როგორც ენდოციტოზის დროს.
პროცესი ცილის სეკრეციანაჩვენებია ნახ. 1.12 მაგრამ.ცილა სინთეზირდება უჯრედის ბირთვთან ახლოს რიბოსომებზე, რომლებიც დაკავშირებულია ენდოპლაზმურ ბადესთან (ე.წ. მარცვლოვანი, ან უხეში, ენდოპლაზმური ბადე); ენდოპლაზმურ რეტიკულუმში მოხვედრისას ცილა იფუთება სატრანსპორტო ვეზიკულებში, რომლებიც გამოყოფილია ორგანელისგან და მიგრირებს გოლჯის აპარატში. აქ ისინი ერწყმის გოლჯის აპარატის ავზებს, სადაც ცილა მოდიფიცირებულია (ე.ი. გარდაიქმნება გლიკოპროტეინად). ცისტერნების ბოლოებზე ვეზიკულები კვლავ გამოეყოფა. სეკრეტორული ვეზიკულები, რომლებიც ატარებენ მოდიფიცირებულ ცილას, მოძრაობენ პლაზმური მემბრანისკენ და ათავისუფლებენ მათ შიგთავსს ეგზოციტოზის გზით.
უჯრედში სატრანსპორტო გზის კიდევ ერთი მაგალითი ნაჩვენებია ნახ. 1.12, B; არის ქოლესტერინის ათვისება უჯრედის მიერ. სისხლში ტრანსპორტირებული ქოლესტერინი ძირითადად ასოცირდება ცილებთან, როგორიცაა ნაწილაკები "დაბალი სიმკვრივის ლიპოპროტეინი"(LNP). ეს ნაწილაკები მემბრანაზე მიმაგრებულია LDL რეცეპტორების სპეციფიკურ უბნებზე, სადაც ხდება ენდოციტოზი და LDL ტრანსპორტირდება უჯრედში "დაფარულ" ვეზიკულებში. ეს ვეზიკულები ერწყმის ენდოსომების წარმოქმნას და ამ პროცესის დროს კარგავენ „ზარს“. თავის მხრივ, ენდოსომები ერწყმის პირველად ლიზოსომებს, რომლებიც შეიცავს უპირატესად ჰიდროლიზურ ფერმენტებს და წარმოქმნიან მეორად, უფრო დიდ ლიზოსომებს. მათში ქოლესტერინი გამოიყოფა LDL ნაწილაკებიდან და დიფუზირდება ციტოზოლში, სადაც ის ხელმისაწვდომი ხდება, მაგალითად, ლიპიდური მემბრანების სინთეზისთვის. ვეზიკულები, რომლებიც არ შეიცავს LDL-ს, ასევე გამოყოფილია ენდოსომებისგან, რომლებიც სპეციალური გზით გადადიან პლაზმურ მემბრანაში და ერწყმის მას, აბრუნებენ მემბრანულ მასალას და, სავარაუდოდ, LDL რეცეპტორებს. იმ მომენტიდან, როდესაც LDL ნაწილაკი მემბრანას უკავშირდება, გადის 10-15 წუთი, სანამ ქოლესტერინი გამოიყოფა მეორადი ლიზოსომიდან. LDL-ის შებოჭვისა და შეწოვის დარღვევა, ანუ უჯრედში ქოლესტერინის მიწოდება, გადამწყვეტ როლს თამაშობს სერიოზული და გავრცელებული დაავადების, ათეროსკლეროზის (არტერიების „გამკვრივება“) განვითარებაში.
არსებობს მრავალი სხვა სატრანსპორტო მარშრუტი, რომელიც მსგავსია ნახ. 1.11 და 1.12,A, რომელთა დახმარებით უჯრედში მოძრაობენ სპეციფიკური ვეზიკულები. ზუსტად არ არის ცნობილი, როგორ მოძრაობენ ისინი, მაგრამ ციტოჩონჩხის ელემენტები, სავარაუდოდ, მონაწილეობენ ამ პროცესში. ვეზიკულებს შეუძლიათ მიკროტუბულების გასწვრივ სრიალება, ამ შემთხვევაში მოძრაობისთვის ენერგია, როგორც ჩანს, უზრუნველყოფილია ვეზიკულასთან ასოცირებული ცილისგან, ATPase (იხ. ქვემოთ). სრულიად გაუგებარი რჩება, რამდენი სხვადასხვა ვეზიკულა, რომლებიც ერთმანეთის მიყოლებით მოძრაობენ ყველა მიმართულებით, მიდიან დანიშნულების ადგილზე. მათ აშკარად უნდა „მონიშვნა“ ისე, რომ სატრანსპორტო სისტემამ აღიაროს და მიზანმიმართულად გადაადგილდეს.
ტრანსპორტირება ორგანელების წარმოქმნით და განადგურებით . აქამდე ჩვენ ვთვლიდით ენდო- და ეგზოციტოზს, როგორც ვეზიკულების შიგთავსის ტრანსპორტირების პროცესებს. არსებობს ამ პროცესების კიდევ ერთი ასპექტი, რომელიც მდგომარეობს იმაში, რომ პლაზმური მემბრანის მიმართული მოცილება უჯრედის ზედაპირის ერთ უბანში ენდოციტოზის გზით და, პირიქით, მისი დამატება მეორეში ეგზოციტოზის გზით, გადაადგილდება უჯრედის მნიშვნელოვანი მონაკვეთები. მემბრანა (ნახ. 1.12.E), რაც უჯრედს აძლევს შესაძლებლობას, მაგალითად, ჩამოაყალიბოს ან გადაადგილდეს.
მსგავსი გადაწყობები ასევე დამახასიათებელია ციტოჩონჩხისთვის, განსაკუთრებით მიკროფილამენტებისა და მიკროტუბულებისთვის (ნახ. 1.1). მიკროფილამენტებიშედგება ძირითადად F-აქტინის ცილარომელსაც შეუძლია ციტოზოლიდან მონომერის პოლიმერიზაციის შედეგად ბოჭკოვან შეკვრაში შეკრება. შეკვრა პოლარიზებულია, ანუ ისინი ხშირად იზრდებიან მხოლოდ ერთი ბოლოდან, აგროვებენ აქტინის ახალ მოლეკულებს, ხოლო მეორე ბოლო ინერტულია ან აქ ხდება დაშლა. ამ პოლარიზებული ზრდის გამო, მიკროფილამენტები ეფექტურად მოძრაობენ და მათი ქსელის სტრუქტურა შეიძლება შეიცვალოს. აქტინის გადასვლა დეპოლიმერიზებული მდგომარეობიდან (სოლ) ორგანიზებულ მდგომარეობაში (გელზე) შეიძლება მოხდეს ძალიან სწრაფად სხვა ცილების გავლენის ქვეშ ან იონის კონცენტრაციის ცვლილებების ქვეშ (იხ. ქვემოთ). ასევე არსებობს ცილები, რომლებიც იწვევენ აქტინის ძაფების მოკლე ფრაგმენტებად დაშლას. მრავალი უჯრედის წვრილი გამონაზარდები - ფილოპოდია - შეიცავს აქტინის ცენტრალურ შეკვრას (ნახ. 1.1), ხოლო ფილოპოდიის სხვადასხვა მოძრაობა, სავარაუდოდ, აქტინური გადასვლებით არის განპირობებული: პოლიმერიზაცია - დეპოლიმერიზაცია.
მიკროტუბულებიასევე ხშირად განიცდიან მსგავს მოძრაობებს. ამ მოძრაობების მექანიზმი მსგავსია - ტუბულინის პოლიმერიზაცია ციტოზოლიდან ისე, რომ მიკროტუბულის ერთი ბოლო იზრდება, მეორე კი ან არ იცვლება, ან იქ ხდება დაშლა. ამრიგად, მიკროტუბულს, მასალის შესაბამისი დამატებით ან მოცილებით, შეუძლია გადაადგილება ციტოზოლში.
ციტოჩონჩხის აქტიური მოძრაობები . ციტოჩონჩხის სტრუქტურებში ცვლილებები შეიძლება მოხდეს როგორც ზემოთ აღწერილი აქტიური მოძრაობების, ისე გადაკეთების შედეგად. ხშირ შემთხვევაში, მიკროტუბულებისა და აქტინის ძაფების მოძრაობა განპირობებულია კონტრაქტული პროტეინებით, რომლებიც აკავშირებენ ძაფებს ან მილაკებს და შეუძლიათ მათი გადაადგილება ერთმანეთთან შედარებით. ციყვები მიოზინი და დინეინიიმყოფება ყველა უჯრედის ციტოზოლში შედარებით მაღალი კონცენტრაციით; ეს არის ელემენტები, რომლებიც ენერგიას გარდაქმნიან მოძრაობად სპეციალიზებულ უჯრედებში (კუნთებში) და ორგანელებში (ცილა). კუნთოვან უჯრედებში მიოზინი აყალიბებს სქელ ძაფებს, რომლებიც ორიენტირებულია აქტინის ძაფების პარალელურად. მიოზინის მოლეკულა თავისი „თავით“ ემაგრება აქტინის ძაფს და ატფ-ის ენერგიის გამოყენებით, ანაცვლებს მიოსინს აქტინის მოლეკულის გასწვრივ.შემდეგ მიოზინი იშლება აქტინისგან. მრავალი ასეთი კავშირი-გათიშვის ციკლის ერთობლიობა იწვევს მაკროსკოპულს კუნთოვანი ბოჭკოების შეკუმშვა(თრ. 4). დინეინი მსგავს როლს ასრულებს მიკროტუბულების მოძრაობაში წამწამების ოპერაციის დროს (ნახ. 1.1). არასპეციალიზებული უჯრედების ციტოპლაზმაში მიოზინი და დინეინი არ ქმნიან რეგულარულ ბოჭკოებს, მაგრამ უმეტეს შემთხვევაში მოლეკულების მცირე ჯგუფებს. ასეთი მცირე აგრეგატების სახითაც კი მათ შეუძლიათ აქტინის ძაფების ან მიკროტუბულების გადაადგილება. ბრინჯი. 1.13 ასახავს ამ პროცესს, როდესაც საპირისპირო პოლარიზებული მიოზინის მოლეკულები ასევე მიმაგრებულია სხვადასხვა მიმართულებით პოლარიზებულ აქტინის ორ ძაფზე. მიოზინის სათავე ჯგუფები იხრება მოლეკულის კუდისკენ, მოიხმარენ ATP-ს, ხოლო აქტინის ორი ძაფი საპირისპირო მიმართულებით ინაცვლებს, რის შემდეგაც მიოზინი იშლება მათგან. ამ ტიპის მოძრაობამ, რომლის დროსაც ATP-ის ენერგია გარდაიქმნება მექანიკურ მუშაობაში, შეუძლია შეცვალოს ციტოჩონჩხის ფორმა და, შესაბამისად, უჯრედები, ასევე უზრუნველყოს ციტოჩონჩხთან დაკავშირებული ორგანელების ტრანსპორტირება.
უჯრედშიდა ტრანსპორტის პროცესები ყველაზე ნათლად შეიძლება გამოვლინდეს ნერვული უჯრედის აქსონზე. აქსონის ტრანსპორტიაქ დეტალურად არის განხილული მოვლენების საილუსტრაციოდ, რომლებიც შესაძლოა მსგავსი გზით მოხდეს უმეტეს უჯრედებში. აქსონი, რომლის დიამეტრი მხოლოდ რამდენიმე მიკრონია, შეიძლება იყოს ერთი მეტრი ან მეტი, და წლები დასჭირდება ცილებს ბირთვიდან აქსონის დისტალურ ბოლოში დიფუზიის გზით გადაადგილებას. დიდი ხანია ცნობილია, რომ როდესაც აქსონის რომელიმე მონაკვეთი განიცდის შეკუმშვას, აქსონის პროქსიმალური ნაწილი ფართოვდება. როგორც ჩანს, ცენტრიდანული ნაკადი დაბლოკილია აქსონში. ასეთი ნაკადი–სწრაფი აქსონის ტრანსპორტირება შეუძლიანაჩვენები იქნება რადიოაქტიური მარკერების მოძრაობით, როგორც ნახ. 1.14. რადიოაქტიურად მარკირებული ლეიცინი შეჰყავდათ ზურგის ფესვის განგლიონში, შემდეგ კი მე-2-დან მე-10 საათამდე რადიოაქტიურობა გაზომეს საჯდომის ნერვში ნეირონების სხეულებიდან 166 მმ მანძილზე. 10 საათის განმავლობაში, ინექციის ადგილზე რადიოაქტიურობის პიკი მნიშვნელოვნად არ შეცვლილა. მაგრამ რადიოაქტიურობის ტალღა გავრცელდა აქსონის გასწვრივ მუდმივი სიჩქარით დაახლოებით 34 მმ 2 საათში, ანუ 410 მმ/დღეში. ნაჩვენებია, რომ ჰომოიოთერმული ცხოველების ყველა ნეირონში, აქსონის სწრაფი ტრანსპორტირება ხდება იმავე სიჩქარით და არ არის შესამჩნევი განსხვავებები თხელ, არამიელინირებულ ბოჭკოებსა და ყველაზე სქელ აქსონებს შორის, აგრეთვე საავტომობილო და სენსორულ ბოჭკოებს შორის. რადიოაქტიური მარკერის ტიპი ასევე არ ახდენს გავლენას სწრაფი აქსონალური ტრანსპორტის სიჩქარეზე; სხვადასხვა რადიოაქტიური ნივთიერებები შეიძლება იყოს მარკერები.

ბრინჯი. 1.13.არაკუნთოვანი მიოზინის კომპლექსი გარკვეული ორიენტაციის მქონე შეიძლება დაუკავშირდეს სხვადასხვა პოლარობის აქტინის ძაფებს და, ატფ-ის ენერგიის გამოყენებით, გადაანაცვლოს ისინი ერთმანეთთან შედარებით.
მოლეკულები, როგორიცაა სხვადასხვა ამინომჟავები, რომლებიც ჩართულია ნეირონის სხეულის პროტეინებში. თუ ჩვენ გავაანალიზებთ ნერვის პერიფერიულ ნაწილს, რათა განვსაზღვროთ აქ გადატანილი რადიოაქტიურობის მატარებლების ბუნება, მაშინ ასეთი მატარებლები გვხვდება ძირითადად ცილოვან ფრაქციაში, არამედ შუამავლების და თავისუფალი ამინომჟავების შემადგენლობაში. იმის ცოდნა, რომ ამ ნივთიერებების თვისებები განსხვავებულია და განსაკუთრებით მათი მოლეკულების ზომები განსხვავებულია, ჩვენ შეგვიძლია ავხსნათ ტრანსპორტის მუდმივი სიჩქარე მხოლოდ ყველა მათგანისთვის საერთო სატრანსპორტო მექანიზმით.
Ზემოთ აღწერილი სწრაფი აქსონის ტრანსპორტიარის ანტეროგრადულიანუ მიმართულია უჯრედის სხეულიდან მოშორებით. ნაჩვენებია, რომ ზოგიერთი ნივთიერება გადადის პერიფერიიდან უჯრედის სხეულში დახმარებით რეტროგრადული ტრანსპორტი.მაგალითად, აცეტილქოლინესტერაზა ამ მიმართულებით ტრანსპორტირდება ორჯერ ნაკლები სიჩქარით, ვიდრე სწრაფი აქსონალური ტრანსპორტის სიჩქარე. მარკერი, რომელიც ხშირად გამოიყენება ნეიროანატომიაში - ხახვი პეროქსიდაზა - ასევე მოძრაობს რეტროგრადულად. რეტროგრადული ტრანსპორტი, ალბათ, მნიშვნელოვან როლს თამაშობს უჯრედულ სხეულში ცილის სინთეზის რეგულირებაში. აქსონის გადაკვეთიდან რამდენიმე დღეში უჯრედის სხეულში შეინიშნება ქრომატოლიზი, რაც ცილის სინთეზის დარღვევაზე მიუთითებს. ქრომატოლიზისთვის საჭირო დრო კორელირებს აქსონის გადაკვეთის ადგილიდან უჯრედის სხეულში რეტროგრადული ტრანსპორტის ხანგრძლივობასთან. ასეთი შედეგი ამ დარღვევის ახსნასაც გვთავაზობს - ირღვევა ცილების სინთეზის მარეგულირებელი „სასიგნალო ნივთიერების“ გადაცემა პერიფერიიდან. ცხადია, ძირითადი "მანქანები" გამოიყენება სწრაფი აქსონისთვის
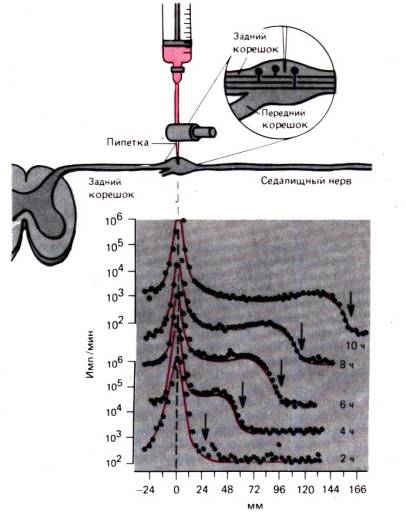
ბრინჯი. 1.14.ექსპერიმენტი, რომელიც აჩვენებს აქსონის სწრაფ ტრანსპორტს კატის საჯდომის ნერვის სენსორულ ბოჭკოებში. ტრიტიირებული ლეიცინი შეჰყავთ დორსალური ფესვის განგლიონში და რადიოაქტიურობა განგლიონსა და სენსორულ ბოჭკოებში იზომება ინექციიდან 2, 4, 6, 8 და 10 საათის შემდეგ. (ფიგურის ქვედა ნაწილი).ავტორი აბსცისამანძილი განგლიონიდან საჯდომის ნერვის მონაკვეთებამდე, სადაც გაზომვა ხდება, გადაიდო. y-ღერძზე, მხოლოდ ზედა და ქვედა მოსახვევებისთვის, რადიოაქტიურობა (imp./min) გამოსახულია ლოგარითმული მასშტაბით. გაზრდილი რადიოაქტიურობის "ტალღა". (ისრები)მოძრაობს 410 მმ/დღეში სიჩქარით
ტრანსპორტი არის ვეზიკულები (ვეზიკულები) და ორგანელები,როგორიცაა ტრანსპორტირებადი ნივთიერებები მიტოქონდრიის შემცველი. ყველაზე დიდი ვეზიკულების ან მიტოქონდრიების მოძრაობა შეიძლება დაფიქსირდეს მიკროსკოპის გამოყენებით in vivo . ასეთი ნაწილაკები აკეთებენ მოკლე, სწრაფ მოძრაობებს ერთ-ერთი მიმართულებით, ჩერდებიან, ხშირად მოძრაობენ ოდნავ უკან ან გვერდზე, ისევ ჩერდებიან და შემდეგ აკეთებენ ჯოხს ძირითადი მიმართულებით. 410 მმ/დღეში შეესაბამება საშუალო ანტეროგრადულ სიჩქარეს დაახლოებით 5 მკმ/წმ; ამიტომ თითოეული ინდივიდუალური მოძრაობის სიჩქარე გაცილებით მაღალი უნდა იყოს და თუ გავითვალისწინებთ ორგანელების, ძაფებისა და მიკროტუბულების ზომას, მაშინ ეს მოძრაობები მართლაც ძალიან სწრაფია. აქსონის სწრაფი ტრანსპორტირება მოითხოვს ატფ-ის მნიშვნელოვან კონცენტრაციას. შხამები, როგორიცაა მიკროტუბულების დამღუპველი კოლხიცინი, ასევე ბლოკავს სწრაფ აქსონალურ ტრანსპორტირებას. აქედან გამომდინარეობს, რომ ტრანსპორტირების პროცესში, რომელსაც განვიხილავთ, ვეზიკულები და ორგანელები მოძრაობენ მიკროტუბულებისა და აქტინის ძაფების გასწვრივ; ეს მოძრაობა უზრუნველყოფილია დინეინისა და მიოზინის მოლეკულების მცირე აგრეგატებით, რომლებიც მოქმედებენ როგორც ნახ. 1.13, ATP-ის ენერგიის გამოყენებით.
აქსონის სწრაფი ტრანსპორტი შეიძლება ასევე იყოს ჩართული პათოლოგიური პროცესები.ზოგიერთი ნეიროტროპული ვირუსი (მაგალითად, ჰერპესი ან პოლიომიელიტის ვირუსები) შეაღწევს აქსონს პერიფერიაზე და რეტროგრადული ტრანსპორტით გადაადგილდება ნეირონების სხეულში, სადაც მრავლდება და ახორციელებს თავის ტოქსიკურ ეფექტს. ტეტანუსის ტოქსინი, ბაქტერიების მიერ წარმოქმნილი ცილა, რომელიც სხეულში შედის კანის დაზიანებების გზით, შეიწოვება ნერვული დაბოლოებით და ტრანსპორტირდება ნეირონულ სხეულში, სადაც იწვევს კუნთების დამახასიათებელ სპაზმს. ცნობილია ტოქსიკური ზემოქმედების შემთხვევები თავად აქსონის ტრანსპორტზე, მაგალითად, სამრეწველო გამხსნელის აკრილამიდის ზემოქმედება. გარდა ამისა, ითვლება, რომ ბერიბერი ბერიბერისა და ალკოჰოლური პოლინეიროპათიის პათოგენეზი მოიცავს სწრაფი აქსონალური ტრანსპორტის დარღვევას.
უჯრედში აქსონის სწრაფი ტრანსპორტის გარდა, ასევე საკმაოდ ინტენსიურია ნელი აქსონის ტრანსპორტი.ტუბულინი მოძრაობს აქსონის გასწვრივ დაახლოებით 1 მმ/დღეში, ხოლო აქტინი უფრო სწრაფად, 5 მმ/დღეში. ციტოჩონჩხის ამ კომპონენტებთან ერთად სხვა ცილებიც მიგრირებენ; მაგალითად, ფერმენტები, როგორც ჩანს, დაკავშირებულია აქტინთან ან ტუბულინთან. ტუბულინისა და აქტინის გადაადგილების სიჩქარე უხეშად შეესაბამება ზრდის ტემპებს, რომლებიც აღმოჩენილია ადრე აღწერილი მექანიზმისთვის, როდესაც მოლეკულები შედის მიკროტუბულის ან მიკროფილამენტის აქტიურ ბოლოში. ამიტომ, ეს მექანიზმი შეიძლება იყოს ნელი აქსონალური ტრანსპორტის საფუძველი. აქსონის ნელი ტრანსპორტირების სიჩქარე ასევე დაახლოებით შეესაბამება აქსონის ზრდის სიჩქარეს, რაც, როგორც ჩანს, მიუთითებს ციტოჩონჩხის სტრუქტურის მიერ დაწესებულ შეზღუდვებზე მეორე პროცესზე.
ამ განყოფილების დასასრულს, ხაზგასმით უნდა აღინიშნოს, რომ უჯრედები არ არის სტატიკური სტრუქტურები, როგორც ჩანს, მაგალითად, ელექტრონულ მიკროსკოპულ ფოტოებში. პლაზმური მემბრანადა განსაკუთრებით ორგანელები მუდმივ სწრაფ მოძრაობაში და მუდმივ რესტრუქტურიზაციაში არიან;ეს არის ერთადერთი მიზეზი, რის გამოც მათ შეუძლიათ ფუნქციონირება. გარდა ამისა, ეს არ არის მარტივი კამერები, რომლებშიც მიმდინარეობს ქიმიური რეაქციები, მაგრამ მემბრანებისა და ბოჭკოების მაღალორგანიზებული კონგლომერატები,რომელშიც რეაქციები მიმდინარეობს ოპტიმალურად ორგანიზებული თანმიმდევრობით.
1.4. უჯრედების ფუნქციების რეგულირება
ცალკეული უჯრედის, როგორც ფუნქციური ერთეულის შენარჩუნება დიდწილად რეგულირდება ბირთვით; ასეთი მარეგულირებელი მექანიზმების შესწავლა უჯრედის ბიოლოგიისა და ბიოქიმიის საგანია. ამავდროულად, უჯრედებმა უნდა შეცვალონ თავიანთი ფუნქციები გარემო პირობებისა და სხეულის სხვა უჯრედების საჭიროებების შესაბამისად, ანუ ისინი ემსახურებიან ფუნქციური რეგულირების ობიექტებს. ქვემოთ მოკლედ განვიხილავთ, როგორ მოქმედებს ეს მარეგულირებელი ზემოქმედება პლაზმურ მემბრანაზე და როგორ აღწევს ისინი უჯრედშიდა ორგანელებში.
მარეგულირებელი ეფექტი უჯრედის მემბრანაზე
მემბრანის პოტენციალი . ხშირ შემთხვევაში, უჯრედული ფუნქციების რეგულირება მემბრანის პოტენციალის შეცვლით ხორციელდება. ლოკალური პოტენციური ცვლილებები შესაძლებელია, როდესაც: 1) მეზობელი უჯრედის არედან ან სხვა უჯრედის მიერ წარმოქმნილი დენი გადის მემბრანაში; 2) იცვლება იონების უჯრედგარე კონცენტრაცია (ხშირად [K + ]გარეთ ); 3) მემბრანის იონური არხები იხსნება. მემბრანის პოტენციალის ცვლილებამ შეიძლება გავლენა მოახდინოს მემბრანის ცილების კონფორმაციაზე, რამაც გამოიწვია, კერძოდ, არხების გახსნა ან დახურვა. როგორც ზემოთ იყო აღწერილი, ზოგიერთი მემბრანული ტუმბოს ფუნქციონირება დამოკიდებულია მემბრანის პოტენციალზე. ნერვული უჯრედები სპეციალიზირებულია მემბრანული პოტენციალის ცვლილებების აღქმაზე, როგორც ინფორმაციას, რომელიც უნდა დამუშავდეს და გადაიცეს (იხ. თავი 2).
უჯრედგარე მარეგულირებელი ნივთიერებები . ყველაზე მნიშვნელოვანი მარეგულირებელი მექანიზმი, რომელიც მოიცავს უჯრედგარე ნივთიერებებს, არის მათი ურთიერთქმედება პლაზმურ მემბრანაზე ან უჯრედის შიგნით არსებულ სპეციფიკურ რეცეპტორებთან. ეს ნივთიერებები მოიცავს სინაფსურ შუამავლებს, რომლებიც გადასცემენ ინფორმაციას ნერვულ უჯრედებს შორის, ადგილობრივ აგენტებსა და ნივთიერებებს შორის, რომლებიც ცირკულირებენ სისხლში და აღწევს სხეულის ყველა უჯრედს, როგორიცაა ჰორმონები და ანტიგენები. სინაფსური ნეიროტრანსმიტერებიარის სინაფსში ნერვული დაბოლოებებიდან გამოთავისუფლებული მცირე მოლეკულები;
როდესაც ისინი მიაღწევენ მეზობელი პოსტსინაფსური უჯრედის პლაზმურ მემბრანას, ისინი იწვევენ ელექტრულ სიგნალებს ან სხვა მარეგულირებელ მექანიზმებს. ეს საკითხი დეტალურად არის განხილული თავში. 3.
ადგილობრივი ქიმიური აგენტები ხშირად გამოიყოფა სპეციალიზებული უჯრედებით. ისინი თავისუფლად იშლება უჯრედგარე სივრცეში, მაგრამ მათი მოქმედება შეზღუდულია უჯრედების მცირე ჯგუფით ამ ნივთიერებების სწრაფი განადგურების გამო, სპონტანურად ან ფერმენტების მოქმედებით. ასეთი აგენტების გათავისუფლების ერთ-ერთი მაგალითია გათავისუფლება ჰისტამინიმასტი უჯრედები დაზიანების ან იმუნური პასუხის დროს. ჰისტამინი იწვევს სისხლძარღვთა გლუვი კუნთების უჯრედების მოდუნებას, ზრდის სისხლძარღვთა ენდოთელიუმის გამტარიანობას და ასტიმულირებს სენსორულ ნერვულ დაბოლოებებს, რომლებიც შუამავლობენ ქავილის შეგრძნებას. სხვა ადგილობრივი ქიმიური აგენტები გამოიყოფა მრავალი სხვა უჯრედის მიერ. ტიპიური ადგილობრივი აგენტები არიან პროსტაგლანდინები,წარმოადგენს დაახლოებით 20 ცხიმოვანი მჟავების წარმოებულების ჯგუფს. ისინი მუდმივად გამოიყოფა ფართოდ განაწილებული უჯრედებიდან, მაგრამ მოქმედებენ მხოლოდ ადგილობრივად, რადგან ისინი სწრაფად ნადგურდებიან მემბრანული ფოსფოლიპაზებით. სხვადასხვა პროსტაგლანდინებს აქვთ მოქმედების ფართო სპექტრი: მათ შეუძლიათ გამოიწვიონ გლუვი კუნთების უჯრედების შეკუმშვა, გამოიწვიოს სისხლის თრომბოციტების (თრომბოციტების) აგრეგაცია ან შეაფერხოს საკვერცხეებში ყვითელი სხეულის განვითარება.
სხვა ადგილობრივი აგენტები ემსახურებიან ზრდის ფაქტორები.ყველაზე ცნობილი ნერვული ზრდის ფაქტორი (NGF) სიმპათიკური ნეირონებისთვის, რომელიც აუცილებელია ამ ნეირონების ზრდისა და გადარჩენისთვის განვითარების დროს. in vivo ან უჯრედულ კულტურაში. ცხადია, ამ კლასის ნეირონების სამიზნე უჯრედები გამოყოფენ NGF და ამით უზრუნველყოფენ სწორ ინერვაციას. ორგანოების ფორმირებისას უჯრედებს ხშირად სჭირდებათ "გამონახონ გზა" სამიზნე უჯრედებისკენ, რომლებიც შეიძლება განლაგდეს მნიშვნელოვან მანძილზე. შესაბამისად, უნდა არსებობდეს მრავალი სპეციალიზებული ზრდის ფაქტორი, როგორიცაა NGF.
ჰორმონები და ანტიგენები სისხლით გადაიტანა ყველა უჯრედში. ანტიგენები იწვევს იმუნურ პასუხს უჯრედებიდან, რომლებიც ატარებენ სპეციფიკურ ანტისხეულებს. თუმცა, ანტიგენები, როგორც წესი, არის უცხო ნივთიერებები, რომლებიც არ წარმოიქმნება რეაქციაში მყოფ ორგანიზმში (დაწვრილებით იხილეთ თავი 18). ზოგიერთი ჰორმონი, როგორიცაა ინსულინი ან თიროქსინი, გავლენას ახდენს უჯრედების მრავალ ტიპზე, ზოგი კი, როგორიცაა სასქესო ჰორმონები, გავლენას ახდენს მხოლოდ უჯრედების გარკვეულ ტიპებზე. ჰორმონები ან პეპტიდებია, რომელთა მოქმედება გამოწვეულია უჯრედის მემბრანის რეცეპტორებთან მათი შეკავშირებით, ან სტეროიდები და თიროქსინი, რომლებიც დიფუზობენ ლიპიდურ მემბრანაში და უკავშირდებიან უჯრედშიდა რეცეპტორებს. სტეროიდული ჰორმონები უკავშირდება ბირთვულ ქრომატინს, რის შედეგადაც ხდება გარკვეული გენების ტრანსკრიფცია. შედეგად წარმოქმნილი ცილები იწვევენ უჯრედული ფუნქციების ცვლილებას, რაც ჰორმონების სპეციფიკური ეფექტია. ჰორმონების გამოყოფასთან და მოქმედებასთან დაკავშირებული საკითხები დეტალურად არის განხილული თავში. 17.
უჯრედშიდა კომუნიკაცია მეორე მესინჯერების მონაწილეობით
ზემოთ აღწერილი მარეგულირებელი ფუნქციები მოიცავს ეფექტებს უჯრედის მემბრანაზე. უჯრედის მემბრანის მიერ მიღებული ინფორმაცია ხშირად უნდა იწვევდეს რეაქციას ორგანელებში და გადაეცემა მათ სხვადასხვა ნივთიერებებით, რომლებიც ცნობილია როგორც მეორე მესინჯერები (პირველისაგან განსხვავებით, უჯრედში გარე წყაროებიდან). მეორე მედიატორების შესწავლა სწრაფად ვითარდება და არ არსებობს იმის გარანტია, რომ პრობლემის დღევანდელი გაგების დონე საკმარისად სრულყოფილი იქნება. აქ შევეხებით სამ კარგად შესწავლილ მედიატორს: Ca 2+, cAMP და ინოზიტოლ ტრიფოსფატს.
კალციუმი.უმარტივესი უჯრედშორისი შუამავალია Ca 2+ იონი. მისი თავისუფალი კონცენტრაცია მოსვენებულ უჯრედში ძალიან დაბალია და შეადგენს 10 -8 -10 -7 მოლ/ლ. მას შეუძლია უჯრედში შევიდეს მემბრანული არხებით, როდესაც ისინი ღიაა, მაგალითად, როდესაც იცვლება მემბრანის პოტენციალი (იხ. თავი 2). Ca 2+-ის შედეგად მიღებული მატება იწვევს უჯრედში მნიშვნელოვან რეაქციებს, როგორიცაა მიოფიბრილების შეკუმშვა, რომელიც არის კუნთების შეკუმშვის საფუძველი (იხ. თავი 4), ან ნეიროტრანსმიტერის შემცველი ვეზიკულების განთავისუფლება ნერვული დაბოლოებებიდან (იხ. თავი 3). . ორივე რეაქცია მოითხოვს Ca 2+ კონცენტრაციას დაახლოებით 10-5 მოლ/ლ. Ca 2+, რომელსაც აქვს მარეგულირებელი ეფექტი, ასევე შეიძლება გამოიყოფა უჯრედშიდა საცავებიდან, როგორიცაა ენდოპლაზმური ბადე. Ca 2+-ის დეპოდან გამოშვება საჭიროებს სხვა შუამავლების მონაწილეობას (იხ. მაგალითად, სურ. 1.16).
ციკლური ადენოზინის მონოფოსფატი, cAMP. ახლახან დადასტურდა, რომ ციკლური ადენოზინ მონოფოსფატი (cAMP), ორგანიზმში ენერგიის მთავარი წყაროს, ატფ-ის წარმოებული, მნიშვნელოვანი მეორე მესინჯერია. რეაქციების რთული ჯაჭვი ნაჩვენებია ნახ. 1.15, იწყება რეცეპტორიდანრ პლაზმური მემბრანის გარე ზედაპირზე, რომელიც შეიძლება იყოს სხვადასხვა შუამავლებისა და ჰორმონების სპეციფიკური შებოჭვის ადგილი. სპეციფიკურ „მასტიმულირებელ“ მოლეკულასთან შეკავშირების შემდეგრ ცვლის კონფორმაციას; ეს ცვლილებები გავლენას ახდენს ცილაზეგს მემბრანის შიდა ზედაპირზე ისე, რომ შესაძლებელი გახდეს ამ უკანასკნელის გააქტიურება უჯრედშიდა გუანოზინტრიფოსფატით (GTP). გააქტიურებული ცილაგს თავის მხრივ, ასტიმულირებს მემბრანის შიდა ზედაპირზე ფერმენტს, ადენილატ ციკლაზას (AC), რომელიც კატალიზებს ატფ-დან cAMP-ის წარმოქმნას. წყალში ხსნადი cAMP და არის შუამავალი, რომელიც გადასცემს ეფექტს

ბრინჯი. 1.15.რეაქციების ჯაჭვი, რომელშიც შედის უჯრედშიდა შუამავალი cAMP (ციკლური ადენოზინმონოფოსფატი). აღმგზნები ან ინჰიბიტორული გარე სიგნალები ააქტიურებენ მემბრანის რეცეპტორებს Rს ან რი . ეს რეცეპტორები არეგულირებენ შებოჭვის პროცესსგ - ცილები უჯრედშიდა GTP-ით (გუანოზინტრიფოსფატი), რითაც ასტიმულირებს ან აინჰიბირებს უჯრედშიდა ადენილატ ციკლაზას (AC). გამაძლიერებელი ფერმენტი AC გარდაქმნის ადენოზინტრიფოსფატს (ATP) cAMP-ად, რომელიც შემდეგ იშლება AMP-მდე ფოსფოდიესტერის დახმარებით. თავისუფალი cAMP დიფუზირდება უჯრედში და ააქტიურებს ადენილატკინაზას (A-კინაზა), ათავისუფლებს მის C კატალიზურ ქვედანაყოფს, რომელიც აკატალიზებს უჯრედშიდა ცილების ფოსფორილირებას, ე.ი. ქმნის უჯრედგარე სტიმულის საბოლოო ეფექტს. სქემა ასევე აჩვენებს ფარმაკოლოგიურ პრეპარატებს და ტოქსინებს, რომლებიც იწვევს (+) ან თრგუნავს (-) ზოგიერთ რეაქციას (მაგრამ ცვლილებებით)
უჯრედგარე რეცეპტორის სტიმულირებარ უჯრედის შიდა სტრუქტურებს.
რეაქციების მასტიმულირებელი ჯაჭვის პარალელურად, რომელიც მოიცავსრ ინჰიბიტორული შუამავლების და ჰორმონების შესაძლო შეკავშირება შესაბამის რეცეპტორთანრ ი რომელიც ისევ GTP-გააქტიურებული პროტეინის მეშვეობითგ აინჰიბირებს AC და, შესაბამისად, cAMP წარმოებას. უჯრედში დიფუზიით, cAMP რეაგირებს ადენილატკინაზასთან (A-კინაზა); ეს ათავისუფლებს C ქვეერთეულს, რომელიც ახდენს ცილის ფოსფორილირების კატალიზებას.ეს ფოსფორილირება აქცევს ცილებს აქტიურ ფორმაში და ახლა მათ შეუძლიათ განახორციელონ თავიანთი სპეციფიკური მარეგულირებელი მოქმედება (მაგალითად, გამოიწვიოს გლიკოგენის დეგრადაცია). ეს რთული მარეგულირებელი სისტემა ძალზე ეფექტურია, რადგან საბოლოო შედეგი არის მრავალი ცილის ფოსფორილირება, ანუ მარეგულირებელი სიგნალი გადის ჯაჭვში დიდი გამაძლიერებელი ფაქტორით. გარე შუამავლები, რომლებიც აკავშირებენ რეცეპტორებს R s და R i თითოეული მათგანისთვის სპეციფიკური ძალიან მრავალფეროვანია. ადრენალინი, არევა R s ან R i მონაწილეობს ლიპიდური და გლიკოგენის მეტაბოლიზმის რეგულაციაში, ასევე გულის კუნთის შეკუმშვაში და სხვა რეაქციებში (იხ. თავი 19). ფარისებრი ჯირკვლის მასტიმულირებელი ჰორმონი, ააქტიურებს რ , ასტიმულირებს ფარისებრი ჯირკვლის მიერ ჰორმონ თიროქსინის გამოყოფას, ხოლო პროსტაგლანდინი I აფერხებს თრომბოციტების აგრეგაციას. ინჰიბიტორული ეფექტები, მათ შორის ადრენალინი, შუამავლობითრ ი გამოხატულია ლიპოლიზის შენელებაში. Ამგვარად, cAMP სისტემა არის მრავალფუნქციური უჯრედშიდა მარეგულირებელი სისტემა,რომლის ზუსტად კონტროლირება შესაძლებელია უჯრედგარე მასტიმულირებელი და ინჰიბიტორული სასიგნალო საშუალებებით.
ინოზიტოლ ფოსფატი "IF თ ". მეორე მედიატორის, ინოზიტოლ ფოსფატის უჯრედშიდა სისტემა სულ ახლახან აღმოაჩინეს (სურ. 1.16). ამ შემთხვევაში, არ არსებობს ინჰიბიტორული გზა, მაგრამ არის მსგავსება cAMP სისტემასთან, რომლის დროსაც R რეცეპტორის სტიმულაციის ეფექტი გადადის მემბრანის შიდა ზედაპირზე GTP-გააქტიურებულ G-პროტეინზე. შემდეგ ეტაპზე, ჩვეულებრივი მემბრანული ლიპიდური ფოსფატიდილინოზიტოლი (PI), რომელმაც ადრე მიიღო ორი დამატებითი ფოსფატური ჯგუფი, გარდაიქმნება PI-დიფოსფატად (FIF 2), რომელიც იხსნება გააქტიურებული ფოსფოდიესტერაზას (PDE) მიერ. ინოზიტოლ ტრიფოსფატი(IFz) და ლიპიდური დიაცილგლიცეროლი(DAG). ინოზიტოლ ტრიფოსფატი არის წყალში ხსნადი მეორე მესინჯერი, რომელიც დიფუზირდება ციტოზოლში. ის ძირითადად მოქმედებს ენდოპლაზმური ბადისგან Ca 2+-ის გამოთავისუფლებით. Ca 2+ თავის მხრივ მოქმედებს როგორც შუამავალი, როგორც ზემოთ იყო აღწერილი; მაგალითად, ის ააქტიურებს Ca2+-დამოკიდებულ ფოსფოკინაზას, რომელიც ფოსფორილირებს ფერმენტებს. DAG-ის ლიპიდური ქვედანაყოფი (ნახ. 1.16) ასევე ატარებს სიგნალს, რომელიც დიფუზირდება პლაზმური მემბრანის ლიპიდურ ფაზაში მის შიდა ზედაპირზე მდებარე C-კინაზამდე, რომელიც აქტიურდება ფოსფატიდილსერინის, როგორც კოფაქტორის მონაწილეობით. შემდეგ C-კინაზა იწვევს ცილების ფოსფორილირებას, გარდაქმნის მათ აქტიურ ფორმაში.
მეორე შუამავალი ინტერფერონ 3-ის უჯრედშიდა სისტემა ასევე შეიძლება კონტროლდებოდეს სხვადასხვა გარეგანი შუამავლებითა და ჰორმონებით, მათ შორის აცეტილქოლინით, სეროტონინით, ვაზოპრესინით და ფარისებრი ჯირკვლის მასტიმულირებელი ჰორმონით; cAMP სისტემის მსგავსად, მას ახასიათებს სხვადასხვა უჯრედშიდა ეფექტი. შესაძლებელია, რომ ეს სისტემა ასევე გააქტიურდეს სინათლის მიერ თვალის ვიზუალურ რეცეპტორში და თამაშობს ცენტრალურ როლს ფოტოტრანსდუქციაში (იხ. თავი 11). პირველად ორგანიზმის ინდივიდუალურ განვითარებაში სპერმატოზოიდით აქტიურდება IGF სისტემის რეცეპტორი, რის შედეგადაც IGF მონაწილეობს კვერცხუჯრედის განაყოფიერების თანმხლებ მარეგულირებელ რეაქციებში.
cAMP და IFz-DAG სისტემები ძალიან ეფექტურია ბიოლოგიური გამაძლიერებლები.Ისინი არიან

ბრინჯი. 1.16.რეაქციების ჯაჭვი, რომელიც მოიცავს უჯრედშიდა შუამავალ ინტერფერონს (ინოზიტოლ ტრიფოსფატი). როგორც cAMP სისტემაში, უჯრედგარე სიგნალი შუამავლობით ხდება ცილის მეშვეობითგ, რომელიც ამ შემთხვევაში ააქტიურებს ფოსფოდიესტერაზას (PDE). ეს ფერმენტი ანგრევს ფოსფატიდილინოზინ დიფოსფატს (FIF) 2 ) პლაზმურ მემბრანაში IF-მდეთ და დიაცილგლიცეროლი (DAG); თუთ დიფუზირდება ციტოპლაზმაში. აქ ის იწვევს Ca-ს გათავისუფლებას 2+ ენდოპლაზმური ბადედან; Ca კონცენტრაციის გაზრდა 2+ ციტოპლაზმაში ([Ca 2+ ]ი ) ააქტიურებს პროტეინ კინაზას, რომელიც ფოსფორილირდება და შესაბამისად ააქტიურებს ფერმენტებს. კიდევ ერთი პროდუქტი, DAG, რჩება მემბრანაში და ააქტიურებს პროტეინ კინაზა C-ს (კოფაქტორ-ფოსფატიდილსერინი, PS). პროტეინ კინაზა C ასევე ფოსფორილირებს ფერმენტებს, რომლებიც შუამავლობენ სპეციფიკურ მოქმედებებს, რომლებიც დაკავშირებულია გარე რეცეპტორის სტიმულაციასთან.რ . რეაქციების ჯაჭვის ტოტები, რომლებიც მოიცავს IFთ და DAG შეიძლება დამოუკიდებლად გააქტიურდეს იონომიცინით და ფორბოლის ეთერით, შესაბამისად (მოდიფიცირებული)
გარდაქმნის რეაქციას შუამავალსა და გარე მემბრანის რეცეპტორს შორის მრავალი უჯრედშიდა ცილის ფოსფორილირებად, რამაც შეიძლება გავლენა მოახდინოს სხვადასხვა უჯრედის ფუნქციებზე. პრობლემის ერთ-ერთი არსებითი ასპექტი ის არის, რომ, რამდენადაც ჩვენ დღეს ვიცით, არსებობს ამ ტიპის მხოლოდ ეს ორი მჭიდროდ დაკავშირებული მარეგულირებელი სისტემა, რომელსაც მრავალი გარე შუამავალი იყენებს სხვადასხვა უჯრედშიდა პროცესების დასარეგულირებლად. ამავდროულად, ეს მარეგულირებელი სისტემები, მათ შორის Ca 2+, მჭიდროდ ურთიერთქმედებენ ერთმანეთთან, რაც მათ საშუალებას აძლევს განახორციელონ უჯრედული ფუნქციების ჯარიმა რეგულირება.
გაკვეთილები და სახელმძღვანელოები
1. ალბერტსი IN., ბრეი დ., ლუის ჯ., რაფ მ., რობერტსი TO., უოტსონ ჯ.დ.უჯრედის მოლეკულური ბიოლოგია, ნიუ-იორკი და ლონდონი, Garland Publishing Inc., 1983 წ.
2. ჩიჰაკ გ., ლონგერ ჰ., ზიგლერ ჰ.(რედ.). ბიოლოგია. ბერლინი, ჰაიდელბერგი, ნიუ-იორკი, სპრინგერი, 1983 წ.
3. ჰილი IN. აგზნებადი მემბრანების იონური არხები. სანდერლენდი, მასაჩუსეტსი, Sinauer Assoc., 1984 წ.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(რედ.). ბიოფიზიკა. ბერლინი, ჰაიდელბერგი, ნიუ-იორკი, სპრინგერი, 1984 წ.
5. იუნგერმანი TO., მალერ ჰ.ბიოქიმია. ბერლინი, ჰაიდელბერგი, ნიუ-იორკი, სპრინგერი, 1980 წ.
6. Kandel E.R., Schwartz-J.H.,(რედ.). ნერვული მეცნიერების პრინციპები, ნიუ-იორკი, ამსტერდამი, ოქსფორდი, ელზევიერი, 1985 წ.
7. შიბლერი თ. ჰ., შმიდტვ.Anatomical des Menschen. ბერლინი, ჰაიდელბერგი, ნიუ-იორკი, ტოკიო, სპრინგერი, 1983 წ.
ორიგინალური სტატიები და მიმოხილვები
8. ბერიჯი მ.ჯ.უჯრედის შიგნით კომუნიკაციის მოლეკულური საფუძველი, მეცნიერება. ამერი 253 124 134 (1985).
9. ბერიჯ მ.ჯ., ირვინ რ.ფ.ინოზიტოლ ტრიფოსფატი, ახალი მეორე მესინჯერი ფიჭური სიგნალის გადაცემაში. Nature, 312, 315 321 (1984).
10. ბრეტშერი მ.ს.უჯრედის მემბრანის მოლეკულები, მეცნიერ. ამერ., 253, 124–134 (1985).
11. დაუთ ჯ.ცოცხალი უჯრედი, როგორც ენერგიის გადამყვანი მანქანა. მიოკარდიუმის მეტაბოლიზმის მინიმალური მოდელი, Biochem. და ბიოფისი. Acta, 895, 41–62 (1987).
12. ჰოჯკინი A.L., Katz IN. ნატრიუმის იონების გავლენა კალმარის გიგანტური აქსონის ელექტრულ აქტივობაზე. ჯ ფიზიოლ. (ლონდონი), 108, 37–77 (1949).
13. ჰოჯკინი A.L., Keynes R.D.კათიონების აქტიური ტრანსპორტი გიგანტურ აქსონებში სეფიადა ლოლიგო,ჯ ფიზიოლ. (ლონდონი), 128, 28–42 (1955).
14. უფრო გრძელი პ.იონური არხები კონფორმაციული სუბსტატიებით, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M.ეგზოპლაზმური ტრანსპორტი ნორმალურ და პათოლოგიურ სისტემებში. In: Physiology and Pathology of Axons, S.G. ცვილის კაცი,რედ. ნიუ-იორკი, Raven Press, 1978 წ.
გეპატიჟებით გაეცნოთ მასალებს და.
: ცელულოზის მემბრანა, მემბრანა, ციტოპლაზმა ორგანელებით, ბირთვი, ვაკუოლები უჯრედის წვენით.პლასტიდების არსებობა მცენარის უჯრედის მთავარი მახასიათებელია.
უჯრედის კედლის ფუნქციები- განსაზღვრავს უჯრედის ფორმას, იცავს გარემო ფაქტორებისგან.
პლაზმური მემბრანა- თხელი ფილმი, შედგება ლიპიდური და ცილის მოლეკულებისგან, ზღუდავს შიდა შიგთავსს გარე გარემოდან, უზრუნველყოფს წყლის, მინერალური და ორგანული ნივთიერებების ტრანსპორტირებას უჯრედში ოსმოსის და აქტიური გადაცემის გზით და ასევე შლის ნარჩენ პროდუქტებს.
ციტოპლაზმა- უჯრედის შიდა ნახევრად თხევადი გარემო, რომელშიც განლაგებულია ბირთვი და ორგანელები, უზრუნველყოფს მათ შორის კავშირებს, მონაწილეობს ცხოვრების ძირითად პროცესებში.
Ენდოპლაზმურ ბადეში- ციტოპლაზმაში განშტოებული არხების ქსელი. მონაწილეობს ცილების, ლიპიდების და ნახშირწყლების სინთეზში, ნივთიერებების ტრანსპორტირებაში. რიბოსომები - სხეულები, რომლებიც მდებარეობს EPS-ზე ან ციტოპლაზმაში, შედგება რნმ-ისა და ცილისგან, მონაწილეობენ ცილის სინთეზში. EPS და რიბოსომები არის ერთი აპარატი ცილების სინთეზისა და ტრანსპორტირებისთვის.
მიტოქონდრია-ორგანელები ციტოპლაზმიდან გამოყოფილია ორი გარსით. მათში ორგანული ნივთიერებები იჟანგება და ატფ-ის მოლეკულები სინთეზირდება ფერმენტების მონაწილეობით. შიდა გარსის ზედაპირის ზრდა, რომელზედაც განლაგებულია ფერმენტები კრისტალების გამო. ATP არის ენერგიით მდიდარი ორგანული ნივთიერება.
პლასტიდები(ქლოროპლასტები, ლეიკოპლასტები, ქრომოპლასტები), მათი შემცველობა უჯრედში მცენარის ორგანიზმის მთავარი მახასიათებელია. ქლოროპლასტები არის პლასტიდები, რომლებიც შეიცავს მწვანე პიგმენტს ქლოროფილს, რომელიც შთანთქავს სინათლის ენერგიას და იყენებს მას ნახშირორჟანგისა და წყლისგან ორგანული ნივთიერებების სინთეზისთვის. ქლოროპლასტების დელიმიტაცია ციტოპლაზმიდან ორი მემბრანით, მრავალი გამონაზარდი - გრანა შიდა მემბრანაზე, რომელშიც ქლოროფილის მოლეკულები და ფერმენტებია განთავსებული.
გოლგის კომპლექსი- ღრუების სისტემა ციტოპლაზმიდან მემბრანით შემოიფარგლება. მათში ცილების, ცხიმებისა და ნახშირწყლების დაგროვება. მემბრანებზე ცხიმებისა და ნახშირწყლების სინთეზის განხორციელება.
ლიზოსომები- ციტოპლაზმიდან ერთი მემბრანით გამოყოფილი სხეულები. მათში შემავალი ფერმენტები აჩქარებენ რთული მოლეკულების მარტივებად დაყოფის რეაქციას: ცილები ამინომჟავებამდე, რთული ნახშირწყლები მარტივზე, ლიპიდები გლიცეროლსა და ცხიმოვან მჟავებამდე და ასევე ანადგურებს უჯრედის მკვდარ ნაწილებს, მთელ უჯრედებს.
ვაკუოლები- უჯრედის წვენით სავსე ციტოპლაზმის ღრუები, სარეზერვო საკვები ნივთიერებების, მავნე ნივთიერებების დაგროვების ადგილი; ისინი არეგულირებენ წყლის შემცველობას უჯრედში.
ბირთვი- უჯრედის ძირითადი ნაწილი, რომელიც გარედან დაფარულია ორმემბრანით, გაჟღენთილია ფორებით ბირთვული გარსით. ნივთიერებები შედიან ბირთვში და გამოიყოფა მისგან ფორების მეშვეობით. ქრომოსომა არის მემკვიდრეობითი ინფორმაციის მატარებლები ორგანიზმის მახასიათებლების, ბირთვის ძირითადი სტრუქტურების შესახებ, რომელთაგან თითოეული შედგება ერთი დნმ-ის მოლეკულისგან ცილებთან ერთად. ბირთვი არის დნმ-ის, i-RNA, r-RNA სინთეზის ადგილი.

გარე მემბრანის არსებობა, ციტოპლაზმა ორგანელებით, ბირთვი ქრომოსომებით.
გარე ან პლაზმური მემბრანა- ზღუდავს უჯრედის შიგთავსს გარემოდან (სხვა უჯრედები, უჯრედშორისი ნივთიერება), შედგება ლიპიდური და ცილის მოლეკულებისგან, უზრუნველყოფს უჯრედებს შორის კომუნიკაციას, ნივთიერებების გადატანას უჯრედში (პინოციტოზი, ფაგოციტოზი) და უჯრედის გარეთ.
ციტოპლაზმა- უჯრედის შიდა ნახევრად თხევადი გარემო, რომელიც უზრუნველყოფს კომუნიკაციას მასში მდებარე ბირთვსა და ორგანელებს შორის. სასიცოცხლო აქტივობის ძირითადი პროცესები მიმდინარეობს ციტოპლაზმაში.
უჯრედის ორგანელები:
1) ენდოპლაზმური რეტიკულუმი (ER)- განშტოებული მილაკების სისტემა, რომელიც მონაწილეობს ცილების, ლიპიდების და ნახშირწყლების სინთეზში, უჯრედში ნივთიერებების ტრანსპორტირებაში;
2) რიბოზომები- rRNA-ს შემცველი სხეულები განლაგებულია ER-ზე და ციტოპლაზმაში და მონაწილეობენ ცილების სინთეზში. EPS და რიბოსომები არის ცილის სინთეზისა და ტრანსპორტირების ერთი აპარატი;
3) მიტოქონდრია- უჯრედის "ელექტროსადგურები", ციტოპლაზმიდან ორი გარსით შემოფარგლული. შინაგანი ქმნის კრისტაებს (ნაკეცებს), რომლებიც ზრდის მის ზედაპირს. კრისტაზე არსებული ფერმენტები აჩქარებენ ორგანული ნივთიერებების დაჟანგვის რეაქციებს და ენერგიით მდიდარი ატფ-ის მოლეკულების სინთეზს;
4) გოლგის კომპლექსი- ციტოპლაზმიდან მემბრანით შემოფარგლული ღრუების ჯგუფი, სავსე ცილებით, ცხიმებითა და ნახშირწყლებით, რომლებიც ან გამოიყენება ცხოვრების პროცესში, ან ამოღებულია უჯრედიდან. კომპლექსის გარსები ახორციელებენ ცხიმებისა და ნახშირწყლების სინთეზს;
5) ლიზოსომები- ფერმენტებით სავსე სხეულები აჩქარებენ ცილების დაშლის რეაქციებს ამინომჟავებამდე, ლიპიდებს გლიცეროლსა და ცხიმოვან მჟავებს, პოლისაქარიდებს მონოსაქარიდებს. ლიზოსომებში, უჯრედის მკვდარი ნაწილები, მთლიანი უჯრედები და უჯრედები განადგურებულია.
უჯრედის ჩანართები- სათადარიგო საკვები ნივთიერებების დაგროვება: ცილები, ცხიმები და ნახშირწყლები.
ბირთვი- უჯრედის ყველაზე მნიშვნელოვანი ნაწილი. იგი დაფარულია ორმემბრანიანი გარსით ფორებით, რომლის მეშვეობითაც ზოგიერთი ნივთიერება აღწევს ბირთვში, ზოგი კი ციტოპლაზმაში. ქრომოსომა არის ბირთვის ძირითადი სტრუქტურები, ორგანიზმის მახასიათებლების შესახებ მემკვიდრეობითი ინფორმაციის მატარებლები. იგი გადაეცემა დედა უჯრედის გაყოფის პროცესში ქალიშვილურ უჯრედებს, ხოლო ჩანასახებთან ერთად - ქალიშვილ ორგანიზმებს. ბირთვი არის დნმ, mRNA, rRNA სინთეზის ადგილი.
Ამოცანა:
ახსენით, რატომ უწოდებენ ორგანელებს უჯრედის სპეციალიზებულ სტრუქტურებს?
პასუხი:ორგანელებს უწოდებენ სპეციალიზებულ უჯრედულ სტრუქტურებს, რადგან ისინი ასრულებენ მკაცრად განსაზღვრულ ფუნქციებს, მემკვიდრეობითი ინფორმაცია ინახება ბირთვში, ATP სინთეზირდება მიტოქონდრიაში, ფოტოსინთეზი მიმდინარეობს ქლოროპლასტებში და ა.
თუ თქვენ გაქვთ შეკითხვები ციტოლოგიასთან დაკავშირებით, შეგიძლიათ დახმარებისთვის მიმართოთ
ევოლუციის მესამე ეტაპი არის უჯრედის გამოჩენა.
ცილების და ნუკლეინის მჟავების მოლეკულები (დნმ და რნმ) ქმნიან ბიოლოგიურ უჯრედს, სიცოცხლის უმცირეს ერთეულს. ბიოლოგიური უჯრედები ყველა ცოცხალი ორგანიზმის „სამშენებლო ბლოკია“ და შეიცავს განვითარების ყველა მატერიალურ კოდს.
დიდი ხნის განმავლობაში მეცნიერები უჯრედის სტრუქტურას უკიდურესად მარტივ თვლიდნენ. საბჭოთა ენციკლოპედიური ლექსიკონი უჯრედის ცნებას ასე განმარტავს: „უჯრედი არის ელემენტარული ცოცხალი სისტემა, ყველა ცხოველისა და მცენარის სტრუქტურისა და სიცოცხლის საფუძველი“. უნდა აღინიშნოს, რომ ტერმინი „დაწყებითი“ არავითარ შემთხვევაში არ ნიშნავს „მარტივს“, პირიქით, უჯრედი არის ღმერთის უნიკალური ფრაქტალური ქმნილება, გასაოცარია თავისი სირთულით და, ამავე დროს, ნაწარმოების განსაკუთრებული თანმიმდევრულობით. მისი ყველა ელემენტიდან.
როდესაც ელექტრონული მიკროსკოპის საშუალებით შევძელით შიგნით შეხედვა, აღმოჩნდა, რომ უმარტივესი უჯრედის მოწყობილობა ისეთივე რთული და გაუგებარია, როგორც თავად სამყარო. დღეს უკვე დადგენილია, რომ "უჯრედი არის სამყაროს განსაკუთრებული მატერია, კოსმოსის განსაკუთრებული მატერია". ერთი უჯრედი შეიცავს ინფორმაციას, რომელიც შეიძლება ჩაიწეროს დიდი საბჭოთა ენციკლოპედიის მხოლოდ რამდენიმე ათეულ ათასობით ტომში. იმათ. უჯრედი, სხვა საკითხებთან ერთად, ინფორმაციის უზარმაზარი „ბიორეზერვუარია“.
მოლეკულური ევოლუციის თანამედროვე თეორიის ავტორი მანფრედ ეიგენი წერს: „იმისათვის, რომ ცილის მოლეკულა შემთხვევით წარმოიქმნას, ბუნებას უნდა გაეკეთებინა დაახლოებით 10130 ცდა და დახარჯოს მოლეკულების ისეთი რაოდენობა, რომელიც საკმარისი იქნება 1027 წლისთვის. სამყაროები. თუ ცილა აგებული იყო ჭკვიანურად, ანუ რომ ყოველი ნაბიჯის მართებულობის შემოწმება შეიძლებოდა რაიმე შერჩევის მექანიზმით, დასჭირდა მხოლოდ 2000 მცდელობა. მივდივართ პარადოქსულ დასკვნამდე: პროგრამა "პრიმიტიული ცოცხალი უჯრედის" მშენებლობისთვის. დაშიფრულია სადღაც ელემენტარული ნაწილაკების დონეზე“.
და სხვანაირად როგორ შეიძლება. თითოეული უჯრედი, რომელსაც აქვს დნმ, დაჯილდოებულია ცნობიერებით, აცნობიერებს საკუთარ თავს და სხვა უჯრედებს და კავშირშია სამყაროსთან, ფაქტობრივად, მისი ნაწილია. და მიუხედავად იმისა, რომ ადამიანის ორგანიზმში უჯრედების რაოდენობა და მრავალფეროვნება გასაოცარია (დაახლოებით 70 ტრილიონი), ისინი ყველა საკუთარი თავის მსგავსია, ისევე როგორც უჯრედებში მიმდინარე ყველა პროცესი ერთნაირია. გერმანელი მეცნიერის როლანდ გლაზერის სიტყვებით, ბიოლოგიური უჯრედების დიზაინი „ძალიან კარგად არის გააზრებული“. ვინ არის კარგად მოფიქრებული?
პასუხი მარტივია: ცილები, ნუკლეინის მჟავები, ცოცხალი უჯრედები და ყველა ბიოლოგიური სისტემა ინტელექტუალური შემოქმედის შემოქმედებითი საქმიანობის პროდუქტია.
რა არის საინტერესო: ატომურ დონეზე, არ არსებობს განსხვავებები ორგანული და არაორგანული სამყაროს ქიმიურ შემადგენლობას შორის. სხვა სიტყვებით რომ ვთქვათ, ატომის დონეზე, უჯრედი იქმნება იგივე ელემენტებისგან, როგორც უსულო ბუნება. განსხვავებები გვხვდება მოლეკულურ დონეზე. ცოცხალ სხეულებში, არაორგანულ ნივთიერებებთან და წყალთან ერთად, ასევე არის ცილები, ნახშირწყლები, ცხიმები, ნუკლეინის მჟავები, ATP სინთეზაზა ფერმენტი და სხვა დაბალი მოლეკულური წონის ორგანული ნაერთები.
დღემდე, უჯრედი ფაქტიურად დაიშალა შესწავლის მიზნით ატომებად. თუმცა, შეუძლებელია მინიმუმ ერთი ცოცხალი უჯრედის შექმნა, რადგან უჯრედის შექმნა ნიშნავს ცოცხალი სამყაროს ნაწილაკების შექმნას. აკადემიკოსი ვ.პ. კაზნაჩეევი თვლის, რომ "უჯრედი არის კოსმოპლანეტარული ორგანიზმი... ადამიანის უჯრედები არის ეთერული ბრუნვის ბიოკოლაიდერების გარკვეული სისტემები. ამ ბიოკოლაიდერებში ხდება ჩვენთვის უცნობი პროცესები, ხდება ნაკადების კოსმოსური ფორმების მატერიალიზაცია, მათი კოსმოსური ტრანსფორმაცია და ამის გამო. ნაწილაკები მატერიალიზდება".
წყალი.
უჯრედის მასის თითქმის 80% წყალია. ბიოლოგიის მეცნიერებათა დოქტორის ს. ზენინის თქმით, წყალი, თავისი კასეტური სტრუქტურიდან გამომდინარე, წარმოადგენს საინფორმაციო მატრიცას ბიოქიმიური პროცესების მართვისთვის. გარდა ამისა, სწორედ წყალია პირველადი „სამიზნე“, რომელთანაც ხმის სიხშირის რხევები ურთიერთქმედებენ. ფიჭური წყლის მოწესრიგება იმდენად მაღალია (ბროლის მოწესრიგებასთან ახლოს), რომ მას თხევად კრისტალს უწოდებენ.
ციყვები.
ცილები მნიშვნელოვან როლს ასრულებენ ბიოლოგიურ ცხოვრებაში. უჯრედი შეიცავს რამდენიმე ათას პროტეინს, რომლებიც უნიკალურია ამ ტიპის უჯრედებისთვის (ღეროვანი უჯრედების გამოკლებით). საკუთარი ცილების სინთეზის უნარი გადაეცემა უჯრედიდან უჯრედს და გრძელდება მთელი სიცოცხლის განმავლობაში. უჯრედის სიცოცხლის განმავლობაში ცილები თანდათან იცვლიან სტრუქტურას, ირღვევა მათი ფუნქცია. ეს დახარჯული ცილები ამოღებულია უჯრედიდან და იცვლება ახლით, რის წყალობითაც უჯრედის სასიცოცხლო აქტივობა შენარჩუნებულია.
უპირველეს ყოვლისა, ჩვენ აღვნიშნავთ ცილების სამშენებლო ფუნქციას, რადგან ისინი წარმოადგენენ სამშენებლო მასალას, რომელიც ქმნის უჯრედების მემბრანებს და უჯრედულ ორგანელებს, სისხლძარღვების კედლებს, მყესებს, ხრტილებს და ა.შ.
ცილების სასიგნალო ფუნქცია ძალზე საინტერესოა. გამოდის, რომ ცილებს შეუძლიათ იმოქმედონ როგორც სასიგნალო ნივთიერებები, გადასცენ სიგნალები ქსოვილებს, უჯრედებსა თუ ორგანიზმებს შორის. სასიგნალო ფუნქციას ასრულებს ჰორმონის პროტეინები. უჯრედებს შეუძლიათ ერთმანეთთან ურთიერთობა დისტანციურად, უჯრედშორისი ნივთიერების მეშვეობით გადაცემული სასიგნალო ცილების გამოყენებით.
ცილებს ასევე აქვთ საავტომობილო ფუნქცია. ყველა სახის მოძრაობა, რომელიც უჯრედებს შეუძლიათ, როგორიცაა კუნთების შეკუმშვა, ხორციელდება სპეციალური კონტრაქტული პროტეინებით. ცილები ასევე ასრულებენ სატრანსპორტო ფუნქციას. მათ შეუძლიათ სხვადასხვა ნივთიერების მიმაგრება და უჯრედის ერთი ადგილიდან მეორეზე გადატანა. მაგალითად, სისხლის ცილა ჰემოგლობინი ანიჭებს ჟანგბადს და ატარებს მას სხეულის ყველა ქსოვილსა და ორგანოში. გარდა ამისა, ცილებს აქვთ დამცავი ფუნქციაც. უცხო ცილების ან უჯრედების ორგანიზმში შეყვანისას მასში წარმოიქმნება სპეციალური ცილები, რომლებიც აკავშირებენ და ანეიტრალებენ უცხო უჯრედებსა და ნივთიერებებს. და ბოლოს, ცილების ენერგეტიკული ფუნქცია არის ის, რომ 1 გ ცილის სრული დაშლით, ენერგია გამოიყოფა 17,6 კჯ ოდენობით.
უჯრედის სტრუქტურა.
უჯრედი შედგება სამი განუყოფლად დაკავშირებული ნაწილისგან: მემბრანა, ციტოპლაზმა და ბირთვი, ხოლო ბირთვის სტრუქტურა და ფუნქცია უჯრედის სიცოცხლის სხვადასხვა პერიოდში განსხვავებულია. უჯრედის სიცოცხლე მოიცავს ორ პერიოდს: გაყოფა, რის შედეგადაც წარმოიქმნება ორი ქალიშვილი უჯრედი და გაყოფებს შორის პერიოდი, რომელსაც ინტერფაზა ეწოდება.
უჯრედის მემბრანა უშუალოდ ურთიერთქმედებს გარე გარემოსთან და ურთიერთქმედებს მეზობელ უჯრედებთან. იგი შედგება გარე ფენისა და პლაზმური მემბრანისგან, რომელიც მდებარეობს მის ქვეშ. ცხოველური უჯრედების ზედაპირულ ფენას გლიკოკალი ეწოდება. ის აკავშირებს უჯრედებს გარე გარემოსთან და მის გარშემო არსებულ ყველა ნივთიერებასთან. მისი სისქე 1 მიკრონზე ნაკლებია.
უჯრედის სტრუქტურა
უჯრედის მემბრანა არის უჯრედის ძალიან მნიშვნელოვანი ნაწილი. იგი აერთიანებს ყველა ფიჭურ კომპონენტს და ზღუდავს გარე და შიდა გარემოს.
უჯრედებსა და გარე გარემოს შორის ხდება ნივთიერებების მუდმივი გაცვლა. წყალი, სხვადასხვა მარილები ცალკეული იონების სახით, არაორგანული და ორგანული მოლეკულები უჯრედში შედიან გარე გარემოდან. ნივთიერებათა ცვლის პროდუქტები, ისევე როგორც უჯრედში სინთეზირებული ნივთიერებები: ცილები, ნახშირწყლები, ჰორმონები, რომლებიც წარმოიქმნება სხვადასხვა ჯირკვლის უჯრედებში, უჯრედიდან მემბრანის მეშვეობით გამოიყოფა გარე გარემოში. ნივთიერებების ტრანსპორტირება პლაზმური მემბრანის ერთ-ერთი მთავარი ფუნქციაა.
ციტოპლაზმა- შიდა ნახევრად თხევადი საშუალება, რომელშიც მიმდინარეობს ძირითადი მეტაბოლური პროცესები. ბოლო კვლევებმა აჩვენა, რომ ციტოპლაზმა არ არის ერთგვარი ხსნარი, რომლის კომპონენტები ურთიერთქმედებენ ერთმანეთთან შემთხვევითი შეჯახებისას. ის შეიძლება შევადაროთ ჟელეს, რომელიც გარე გავლენის საპასუხოდ იწყებს „კანკალს“. ასე აღიქვამს და გადასცემს ინფორმაციას ციტოპლაზმა.
ბირთვი და სხვადასხვა ორგანელები განლაგებულია ციტოპლაზმაში, რომლებიც გაერთიანებულია მის მიერ ერთ მთლიანობაში, რაც უზრუნველყოფს მათ ურთიერთქმედებას და უჯრედის, როგორც ერთიანი ინტეგრალური სისტემის აქტივობას. ბირთვი მდებარეობს ციტოპლაზმის ცენტრალურ ნაწილში. ციტოპლაზმის მთელი შიდა ზონა ივსება ენდოპლაზმური რეტიკულუმით, რომელიც წარმოადგენს უჯრედულ ორგანოიდს: გარსებით შემოსაზღვრული მილაკების, ვეზიკულებისა და „ცისტერნების“ სისტემა. ენდოპლაზმური ბადე მონაწილეობს მეტაბოლურ პროცესებში, უზრუნველყოფს ნივთიერებების ტრანსპორტირებას გარემოდან ციტოპლაზმაში და ცალკეულ უჯრედშიდა სტრუქტურებს შორის, მაგრამ მისი მთავარი ფუნქციაა ცილის სინთეზში მონაწილეობა, რომელიც ხორციელდება რიბოსომებში. - მრგვალი ფორმის მიკროსკოპული პატარა სხეულები 15-20 ნმ დიამეტრით. სინთეზირებული ცილები ჯერ გროვდება ენდოპლაზმური რეტიკულუმის არხებსა და ღრუებში და შემდეგ ტრანსპორტირდება ორგანელებსა და უჯრედულ უბნებში, სადაც ისინი მოიხმარენ.
ციტოპლაზმაში ცილებთან ერთად ასევე არის მიტოქონდრიები, 0,2-7 მიკრონი ზომის პატარა სხეულები, რომლებსაც უჯრედების „ელექტროსადგურებს“ უწოდებენ. რედოქს რეაქციები ხდება მიტოქონდრიებში, რაც უჯრედებს ენერგიით უზრუნველყოფს. მიტოქონდრიების რაოდენობა ერთ უჯრედში მერყეობს რამდენიმედან რამდენიმე ათასამდე.
ბირთვი- უჯრედის სასიცოცხლო მნიშვნელობის ნაწილი, აკონტროლებს ცილების სინთეზს და მათი მეშვეობით უჯრედში მიმდინარე ყველა ფიზიოლოგიურ პროცესს. განუყოფელი უჯრედის ბირთვში განასხვავებენ ბირთვულ მემბრანას, ბირთვის წვენს, ბირთვს და ქრომოსომებს. ბირთვული კონვერტის საშუალებით ხდება ნივთიერებების უწყვეტი გაცვლა ბირთვსა და ციტოპლაზმას შორის. ბირთვული კონვერტის ქვეშ - ბირთვული წვენი (ნახევრად თხევადი ნივთიერება), რომელიც შეიცავს ბირთვს და ქრომოსომებს. ნუკლეოლი არის მკვრივი მომრგვალებული სხეული, რომლის ზომები შეიძლება განსხვავდებოდეს ფართოდ, 1-დან 10 მიკრონიმდე და მეტი. იგი ძირითადად შედგება რიბონუკლეოპროტეინებისგან; მონაწილეობს რიბოზომების წარმოქმნაში. ჩვეულებრივ უჯრედში არის 1-3 ბირთვი, ზოგჯერ რამდენიმე ასეულამდე. ბირთვი შედგება რნმ-ისა და ცილისგან.
უჯრედის მოსვლასთან ერთად სიცოცხლე გაჩნდა დედამიწაზე!
Გაგრძელება იქნება...

