Biztosítja a sejt és a környezet kapcsolatát. A szervezet kapcsolata a környezettel
Egy szervezet kapcsolata a környezettel fizikai-kémiai szempontból nyitott rendszer, vagyis olyan rendszer, ahol biokémiai folyamatok folynak. A kiindulási anyagok a környezetből származnak, a szintén folyamatosan képződő anyagok pedig kikerülnek. A szervezetben a többirányú reakciók termékeinek sebessége és koncentrációja közötti egyensúly feltételes, képzeletbeli, mivel az anyagok felvétele és eltávolítása nem áll meg. Folyamatos kapcsolat a környezettel, és lehetővé teszi számunkra, hogy egy élő szervezetet nyitott rendszernek tekintsünk.
A Nap minden élő sejt energiaforrása. A növényi sejtek klorofill segítségével rögzítik a napfény energiáját, és a fotoszintézis során asszimilációs reakciókhoz használják fel. Az állatok, gombák, baktériumok sejtjei közvetetten használják fel a napenergiát, amikor egy földi növény által szintetizált szerves anyagokat szétosztják.
A sejt tápanyagainak egy része a sejtlégzés során lebomlik, így biztosítva a különféle sejttevékenységekhez szükséges energiát. Ez a folyamat a mitokondriumoknak nevezett organellumokban játszódik le. A mitokondrium két membránból áll: a külső, amely elválasztja az organellumát a citoplazmától, és a belső, amely számos redőt alkot. A légzés fő terméke az ATP. Kilép a mitokondriumokból, és energiaforrásként szolgál számos kémiai reakcióhoz a citoplazmában és a sejtmembránban. Ha a sejtlégzés megvalósításához oxigénre van szükség, akkor a légzést aerob légzésnek nevezzük, de ha a reakciók oxigén hiányában mennek végbe, akkor anaerob légzésről beszélünk.
A sejtben végzett bármilyen munkához az energiát egyetlen formában használják fel - az ATP foszfátkötéseiből származó energia formájában. Az ATP egy nagyon mobil vegyület. Az ATP képződése a mitokondriumok belső membránján történik. Az ATP szintetizálódik minden sejtben a légzés során a szénhidrátok, zsírok és más szerves anyagok oxidációs energiája miatt. A zöld növények sejtjeiben az ATP fő mennyisége a kloroplasztiszokban szintetizálódik a napenergia hatására. Bennük a fotoszintézis során sokszor több ATP termelődik, mint a mitokondriumokban. Az ATP a foszfor-oxigén kötések felszakadásával és energia felszabadulásával bomlik le. Ez az ATPáz enzim hatására történik az ATP hidrolízis folyamatában - víz hozzáadásával egy foszforsav molekula eltávolításával. Ennek eredményeként az ATP ADP-vé alakul, és ha két foszforsavmolekula leszakad, akkor AMP-vé. Mindegyik gramm savmolekula hasítási reakcióját 40 kJ felszabadulás kíséri. Ez nagyon nagy energiahozam, ezért az ATP foszfor-oxigén kötéseit általában makroerg (nagy energiájú) kötésnek nevezik.
Az ATP felhasználása a képlékeny csere reakcióiban az ATP hidrolízisével való konjugálással valósul meg. Különböző anyagok molekulái energiával töltődnek fel az ATP molekulából a hidrolízis során felszabaduló foszforcsoport megkötésével, azaz foszforilációval.
A foszfátszármazékok sajátossága, hogy nem hagyják el a sejtet, bár „kisütött” formáik szabadon áthaladnak a membránon. Ennek köszönhetően a foszforilált molekulák a sejtben maradnak mindaddig, amíg a megfelelő reakciókban fel nem használják őket.
Az ADP ATP-vé alakításának fordított folyamata egy foszforsavmolekula ADP-hez kapcsolódásával történik, amely vizet szabadít fel és nagy mennyiségű energiát vesz fel.
Így az ATP univerzális és azonnali energiaforrás a sejttevékenységhez. Ez egyetlen sejtes energiaalapot hoz létre, és lehetővé teszi annak újraelosztását és szállítását a sejt egyik részéből a másikba.
A foszfátcsoport átvitele fontos szerepet játszik a kémiai reakciókban, például a makromolekulák monomerekből történő összeállításában. Például aminosavak csak akkor kombinálhatók peptidekké, ha azokat előzőleg foszforilezték. A mechanikai összehúzódási vagy mozgási folyamatok, az oldott anyag koncentrációgradiens ellenében történő átvitele és más folyamatok az ATP-ben tárolt energiafelhasználással járnak.
Az energiacsere folyamata a következőképpen ábrázolható. A citoplazmában található nagy molekulatömegű szerves anyagok enzimatikusan, hidrolízissel egyszerűbbekké alakulnak, amelyekből állnak: fehérjék - aminosavakká, poli- és diszacharidok - monoszacharidokká (+ glükóz), zsírok glicerinné és zsírsavakká. Az oxidatív folyamatok hiányoznak, kevés energia szabadul fel, ami nem kerül felhasználásra és termikus formába kerül. A legtöbb sejt először szénhidrátot használ. A poliszacharidok (növényekben keményítő, állatokban glikogén) glükózzá hidrolizálódnak. A glükóz oxidációja három fázisban megy végbe: glikolízis, oxidatív dekarboxiláció (Krebs-ciklus – citromsavciklus) és oxidatív foszforiláció (légzési lánc). A citoplazmában a glikolízis, amelynek eredményeként egy glükózmolekula két piroszőlősavmolekulára hasad, két molekula ATP felszabadulásával. Oxigén hiányában a piroszőlősav etanollá (fermentáció) vagy tejsavvá (anaerob légzés) alakul.
Ha a glikolízist állati sejtekben végzik, a hat szénatomos glükózmolekula két tejsavmolekulára bomlik. Ez a folyamat többlépcsős. Ezt egymás után 13 enzim hajtja végre. Az alkoholos fermentáció során egy glükózmolekulából két molekula etanol és két molekula CO2 képződik.
A glikolízis az anaerob és aerob légzés közös fázisa, a másik kettőt csak aerob körülmények között hajtják végre. Az oxigénmentes oxidáció folyamata, amelyben a metabolitok energiájának csak egy része szabadul fel és kerül felhasználásra, a végső folyamat az anaerob szervezetek számára. Oxigén jelenlétében a piroszőlősav átjut a mitokondriumokba, ahol számos egymást követő reakció eredményeként teljesen oxidálódik aerob módon H2O-vá és CO2-vé, miközben az ADP egyidejűleg foszforilálódik ATP-vé. Ugyanakkor a glikolízis két ATP-molekulát ad, kettőt - a Krebs-ciklust, 34 - a légzőláncot. Egy glükózmolekula H2O-vá és CO2-vé történő teljes oxidációjából származó nettó hozam 38 molekula.
Így az aerob szervezetekben a szerves anyagok végső lebontása légköri oxigénnel egyszerű szervetlen anyagokká: CO2 és H2O oxidálásával történik. Ez a folyamat a mitokondriumok krisztjain megy végbe. Ilyenkor a maximális mennyiségű szabad energia szabadul fel, melynek jelentős része az ATP molekulákban van lefoglalva. Könnyen belátható, hogy az aerob oxidáció biztosítja a sejt számára a legnagyobb mértékben a szabad energiát.
A katabolizmus hatására a sejtben energiadús ATP molekulák halmozódnak fel, CO2 és felesleges víz kerül a külső környezetbe.
A légzéshez nem szükséges cukormolekulák a sejtben tárolhatók. A felesleges lipidek vagy lehasadnak, majd ezek hasítási termékei a légzés szubsztrátjaként a mitokondriumokba kerülnek, vagy zsírcseppek formájában tartalékba rakódnak a citoplazmában. A fehérjék a sejtbe bejutó aminosavakból épülnek fel. A fehérjeszintézis a riboszómáknak nevezett organellumokban megy végbe. Minden riboszóma két részrészecskéből áll – nagy és kicsi: mindkét részecske tartalmaz fehérje- és RNS-molekulákat.
A riboszómák gyakran egy speciális membránrendszerhez kapcsolódnak, amely tartályokból és hólyagokból áll, az úgynevezett endoplazmatikus retikulumhoz (ER); a sok fehérjét termelő sejtekben az endoplazmatikus retikulum gyakran nagyon jól fejlett és tele van riboszómákkal. Egyes enzimek csak akkor hatásosak, ha egy membránhoz kapcsolódnak. A lipidszintézisben részt vevő enzimek többsége itt található. Így az endoplazmatikus retikulum egyfajta sejtasztal.
Ezenkívül az ER a citoplazmát külön szakaszokra vagy kompartmentekre bontja, azaz szétválasztja a citoplazmában egyidejűleg előforduló különféle kémiai folyamatokat, és ezáltal csökkenti annak valószínűségét, hogy ezek a folyamatok interferálják egymást.
Az adott sejt által képződött termékek gyakran a sejten kívül kerülnek felhasználásra. Ilyen esetekben a riboszómákon szintetizált fehérjék áthaladnak az endoplazmatikus retikulum membránjain, és a körülöttük kialakuló membránvezikulákba csomagolódnak, amelyeket aztán az ER-ből kifűznek. Ezek a buborékok ellaposodva és egymásra halmozódva, mint egy kupacba rakott palacsinta, jellegzetes szerkezetet alkotnak, amelyet Golgi-komplexumnak vagy Golgi-apparátusnak neveznek. A Golgi-készülékben való tartózkodásuk során a fehérjék bizonyos változásokon mennek keresztül. Amikor eljön az ideje, hogy elhagyják a sejtet, a hártyás vezikulák összeolvadnak a sejtmembránnal és kiürülnek, tartalmukat kifelé öntik, azaz exocitózissal történik a szekréció.
A Golgi-készülékben lizoszómák is képződnek - emésztőenzimeket tartalmazó membrántasakok. A modern citológia egyik leglenyűgözőbb ága annak megértése, hogy egy sejt hogyan állít elő, csomagol és exportál bizonyos fehérjéket, és hogyan „tudja”, hogy mely fehérjéket kell megtartania magának.
Bármely sejt membránja folyamatosan mozog és változik. Az ER membránok lassan mozognak a sejtben. E membránok külön szakaszai elkülönülnek, és hólyagokat képeznek, amelyek átmenetileg a Golgi-készülék részévé válnak, majd az exocitózis folyamatában egyesülnek a sejtmembránnal.
Később a membránanyag visszakerül a citoplazmába, ahol újra felhasználják.
A sejt külső membránján keresztül történik a sejtbe jutó, vagy általa kibocsátott anyagok kifelé irányuló cseréje, valamint a különböző jelek cseréje a mikro- és makrokörnyezettel. Mint ismeretes, a sejtmembrán egy lipid kettős réteg, amelybe különböző fehérjemolekulák vannak beágyazva, amelyek speciális receptorokként, ioncsatornákként, különféle vegyi anyagokat aktívan továbbító vagy eltávolító eszközökként, sejtközi érintkezésekként stb. a membrán aszimmetrikusan: külső felülete szfingomielinből és foszfatidilkolinból, belső felülete foszfatidil-szerinből és foszfatidil-etanol-aminból áll. Az ilyen aszimmetria fenntartása energiaráfordítást igényel. Ezért a sejt károsodása, fertőzése, energiaéhezése esetén a membrán külső felülete számára szokatlan foszfolipidekkel gazdagodik, amely megfelelő reakcióval a sejtkárosodás jelzésévé válik más sejtek és enzimek számára. A legfontosabb szerepet a foszfolipáz A2 oldható formája tölti be, amely az arachidonsavat lebontja, és a fenti foszfolipidekből lizoformákat hoz létre. Az arachidonsav korlátozó láncszem az olyan gyulladásos mediátorok létrehozásában, mint az eikozanoidok, és a védőmolekulák - pentraxinok (C-reaktív fehérje (CRP), amiloid fehérjék prekurzorai) - kapcsolódnak a membrán lizoformáihoz, amit a membrán aktiválása követ. komplement rendszer a klasszikus útvonal és a sejtpusztulás mentén.
A membrán szerkezete hozzájárul a sejt belső környezete jellemzőinek, a külső környezettől való eltéréseinek megőrzéséhez. Ezt a sejtmembrán szelektív permeabilitása, a benne lévő aktív transzportmechanizmusok megléte biztosítja. Megsértésük közvetlen károsodás következtében, például tetrodotoxin, ouabain, tetraetil-ammónium, vagy a megfelelő „szivattyúk” elégtelen energiaellátása esetén a sejt elektrolit-összetételének megsértéséhez, metabolizmusának megváltozásához vezet. , bizonyos funkciók megsértése - összehúzódás, gerjesztő impulzus vezetése stb. A sejt ioncsatornáinak (kalcium, nátrium, kálium és klorid) megsértése emberben genetikailag is meghatározható az ezek szerkezetéért felelős gének mutációjával csatornák. Az úgynevezett channelopathiák az ideg-, izom- és emésztőrendszer örökletes betegségeinek okozói. A sejten belüli túlzott vízbevitel a komplement aktiválása során bekövetkező membránperforáció vagy a citotoxikus limfociták és természetes gyilkosok támadása miatt annak repedéséhez - citolízishez - vezethet.
A sejtmembránba sok receptor épül be – olyan struktúrák, amelyek a megfelelő specifikus jelmolekulákkal (ligandumokkal) kombinálva jelet továbbítanak a sejtbe. Ez különböző szabályozó kaszkádokon keresztül történik, amelyek enzimatikusan aktív molekulákból állnak, amelyek szekvenciálisan aktiválódnak, és végső soron hozzájárulnak a különböző sejtprogramok végrehajtásához, mint például a növekedés és proliferáció, a differenciálódás, a motilitás, az öregedés és a sejthalál. A szabályozási kaszkádok meglehetősen sokak, de számukat még nem határozták meg teljesen. A sejten belül is létezik a receptorok rendszere és a hozzájuk kapcsolódó szabályozó kaszkádok; egy bizonyos szabályozó hálózatot hoznak létre a sejt funkcionális állapotától, fejlődési stádiumától és más receptoroktól érkező jelek egyidejű hatásától függően a koncentráció, eloszlás és a további jelút megválasztásával. Ennek eredménye lehet a jel gátlása vagy felerősítése, más szabályozási útvonalon történő iránya. Mind a receptor apparátus, mind pedig a szabályozó kaszkádokon, például a sejtmag felé vezető jelátviteli útvonalak megszakadhatnak egy genetikai hiba következtében, amely a szervezet szintjén veleszületett hibaként vagy egy adott sejt szomatikus mutációja miatt következik be. típus. Ezeket a mechanizmusokat károsíthatják a fertőző ágensek, méreganyagok, és az öregedés során is megváltozhatnak. Ennek végső szakasza lehet a sejt funkcióinak, szaporodási és differenciálódási folyamatainak megsértése.
Az intercelluláris kölcsönhatás folyamataiban fontos szerepet játszó molekulák is a sejtek felszínén helyezkednek el. Ezek lehetnek sejtadhéziós fehérjék, szöveti kompatibilitási antigének, szövetspecifikus, differenciáló antigének stb. Ezeknek a molekuláknak az összetételében bekövetkező változások az intercelluláris kölcsönhatások megsértését idézhetik elő, és aktiválhatják az ilyen sejtek eliminációjához szükséges megfelelő mechanizmusokat. bizonyos veszélyt jelentenek a szervezet integritására, mint a fertőzések, különösen a vírusok, tározója, vagy mint a tumor növekedésének potenciális elindítói.
A sejt energiaellátásának megsértése
A sejt energiaforrása a táplálék, melynek lebomlása után energia szabadul fel a végső anyagokba. A mitokondriumok az energiatermelés fő helyei, amelyekben az anyagok a légzőlánc enzimei segítségével oxidálódnak. Az oxidáció a fő energiaszolgáltató, mivel a glikolízis eredményeként az oxidációhoz képest ugyanannyi oxidációs szubsztrátumból (glükóz) legfeljebb 5% energia szabadul fel. Az oxidáció során felszabaduló energia mintegy 60%-a oxidatív foszforilációval halmozódik fel makroerg foszfátokban (ATP, kreatin-foszfát), a többi hő formájában disszipálódik. A jövőben a nagyenergiájú foszfátokat a sejt olyan folyamatokhoz használja fel, mint a szivattyúzás, szintézis, osztódás, mozgás, szekréció stb. Három olyan mechanizmus létezik, amelyek károsodása zavart okozhat a sejt energiaellátásában: az első az energia-anyagcsere enzimek szintézisének mechanizmusa, a második az oxidatív foszforiláció mechanizmusa, a harmadik az energiafelhasználás mechanizmusa.
Az elektrontranszport megsértése a mitokondriumok légzési láncában vagy az ADP oxidáció és foszforiláció szétkapcsolása a protonpotenciál elvesztésével - az ATP képződés hajtóereje - az oxidatív foszforiláció gyengüléséhez vezet, oly módon, hogy az energia nagy része a szervezetben disszipálódik. csökken a hőforma és a makroerg vegyületek száma. Az adrenalin hatására létrejövő oxidáció és foszforiláció szétkapcsolását a homoioterm organizmusok sejtjei a hőtermelés fokozására használják, miközben állandó testhőmérsékletet tartanak fenn lehűléskor, illetve annak emelkedése során. A thyreotoxicosisban jelentős változások figyelhetők meg a mitokondriumok szerkezetében és az energiaanyagcserében. Ezek a változások kezdetben reverzibilisek, de egy bizonyos pont után visszafordíthatatlanná válnak: a mitokondriumok széttöredeznek, szétesnek vagy megduzzadnak, krisztusokat veszítenek, vakuolákká alakulnak, és végül olyan anyagokat halmoznak fel, mint a hialin, ferritin, kalcium, lipofuscin. A skorbutban szenvedő betegeknél a mitokondriumok összeolvadnak, és kondrioszférákat képeznek, valószínűleg a peroxidvegyületek által okozott membránkárosodás miatt. A mitokondriumok jelentős károsodása az ionizáló sugárzás hatására, a normál sejt rosszindulatúvá történő átalakulása során következik be.
A mitokondriumok a kalciumionok erőteljes depója, ahol koncentrációja több nagyságrenddel magasabb, mint a citoplazmában. Ha a mitokondriumok károsodnak, a kalcium bejut a citoplazmába, ami a proteinázok aktiválódását okozza az intracelluláris struktúrák károsodásával és a megfelelő sejt funkcióinak megzavarásával, például kalcium-kontraktúrákkal vagy akár „kalciumhalál”-val az idegsejtekben. A mitokondriumok funkcionális képességének megsértése következtében a szabad gyökös peroxidvegyületek képződése élesen megnövekszik, amelyek nagyon magas reakcióképességgel rendelkeznek, és ezért károsítják a fontos sejtkomponenseket - nukleinsavakat, fehérjéket és lipideket. Ez a jelenség az úgynevezett oxidatív stressz során figyelhető meg, és negatív következményekkel járhat a sejt létére nézve. Így a külső mitokondriális membrán károsodása a membránközi térben található anyagok, elsősorban a citokróm C és néhány más biológiailag aktív anyag citoplazmába történő felszabadulásával jár, amelyek programozott sejthalált - apoptózist - okozó láncreakciókat váltanak ki. A mitokondriális DNS károsításával a szabadgyök-reakciók eltorzítják az egyes légzési lánc enzimek kialakulásához szükséges genetikai információkat, amelyek kifejezetten a mitokondriumokban termelődnek. Ez az oxidatív folyamatok még nagyobb megzavarásához vezet. Összességében a mitokondriumok belső genetikai apparátusa a sejtmag genetikai apparátusához képest kevésbé védett a benne kódolt genetikai információt megváltoztatni képes káros hatásokkal szemben. Ennek eredményeként a mitokondriális diszfunkció az élet során előfordul, például az öregedés folyamatában, a sejt rosszindulatú átalakulása során, valamint a tojásban lévő mitokondriális DNS mutációjával kapcsolatos örökletes mitokondriális betegségek hátterében. Jelenleg több mint 50 mitokondriális mutációt írtak le, amelyek az ideg- és izomrendszer örökletes degeneratív betegségeit okozzák. Kizárólag az anyától továbbítják őket a gyermeknek, mivel a spermium mitokondriumai nem részei a zigótának, és ennek megfelelően az új szervezetnek.
A genetikai információ megőrzésének és továbbításának megsértése
A sejtmag tartalmazza a legtöbb genetikai információt, és így biztosítja normális működését. A szelektív génexpresszió segítségével interfázisban koordinálja a sejt munkáját, tárolja a genetikai információkat, újrateremti és átadja a genetikai anyagot a sejtosztódás folyamatában. A DNS-replikáció és az RNS-transzkripció a sejtmagban megy végbe. Különféle patogén tényezők, például ultraibolya és ionizáló sugárzás, szabad gyökök oxidációja, vegyszerek, vírusok károsíthatják a DNS-t. Becslések szerint egy melegvérű állat minden sejtje 1 nap alatt. több mint 10 000 bázist veszít. Ehhez hozzá kell adni a megsértéseket az osztás közbeni másoláskor. Ha ez a károsodás továbbra is fennáll, a sejt nem tudna túlélni. A védelem az olyan erős javítórendszerek létezésében rejlik, mint az ultraibolya endonukleáz, a reparatív replikáció és a rekombinációs javítás rendszere, amelyek helyettesítik a DNS-károsodást. A reparatív rendszerek genetikai hibái betegségek kialakulását okozzák a DNS-károsító tényezők iránti fokozott érzékenység miatt. Ez egy pigmentált xeroderma, valamint néhány felgyorsult öregedési szindróma, amelyet a rosszindulatú daganatok megjelenésére való fokozott hajlam kísér.
A DNS-replikáció folyamatainak szabályozási rendszere, a hírvivő RNS (mRNS) transzkripciója, a genetikai információ nukleinsavakból a fehérjék szerkezetébe történő transzlációja meglehetősen összetett és többszintű. A több mint 3000, bizonyos géneket aktiváló transzkripciós faktor hatását kiváltó szabályozó kaszkádokon kívül létezik egy kis RNS-molekulák (interferáló RNS-ek; RNAi) által közvetített többszintű szabályozórendszer is. A körülbelül 3 milliárd purin és pirimidin bázisból álló emberi genom a fehérjeszintézisért felelős szerkezeti gének mindössze 2%-át tartalmazza. A többi a szabályozó RNS-ek szintézisét biztosítja, amelyek a transzkripciós faktorokkal együtt a kromoszómák DNS-szintjén aktiválják vagy blokkolják a szerkezeti gének munkáját, vagy befolyásolják a hírvivő RNS (mRNS) transzlációját a citoplazmában egy polipeptid molekula képződése során. . A genetikai információ megsértése előfordulhat mind a strukturális gének szintjén, mind a DNS szabályozó részének megfelelő megnyilvánulásaival különböző örökletes betegségek formájában.
A közelmúltban nagy figyelmet fordítanak a genetikai anyag azon változásaira, amelyek egy organizmus egyedfejlődése során következnek be, és amelyek a DNS és a kromoszómák egyes szakaszainak gátlásával vagy aktiválásával járnak, ezek metilációja, acetilezése és foszforilációja következtében. Ezek a változások hosszú ideig fennállnak, néha a szervezet egész életében az embriogenezistől az idős korig, és epigenomikus öröklődésnek nevezik.
A megváltozott genetikai információval rendelkező sejtek szaporodását a mitotikus ciklus szabályozási rendszerei (tényezői) is megakadályozzák. Kölcsönhatásba lépnek ciklinfüggő protein kinázokkal és katalitikus alegységeikkel - ciklinekkel -, és blokkolják a teljes mitotikus ciklus áthaladását a sejtben, leállítva az osztódást a preszintetikus és szintetikus fázis határán (G1/S blokk), amíg a DNS-javítás be nem fejeződik. , és ha ez lehetetlen, beindítják a programozott halálsejteket. E tényezők közé tartozik a p53 gén, amelynek mutációja a transzformált sejtek szaporodása feletti kontroll elvesztését okozza; az emberi rákos megbetegedések csaknem 50%-ában fordul elő. A mitotikus ciklus áthaladásának második ellenőrző pontja a G2/M határon található. Itt a kromoszómaanyag helyes eloszlását a mitózisban vagy meiózisban lévő leánysejtek között a sejtorsót, a központot és a centromereket (kinetokorokat) szabályozó mechanizmusok komplexével szabályozzák. E mechanizmusok hatástalansága a kromoszómák vagy részeik eloszlásának megsértéséhez vezet, ami abban nyilvánul meg, hogy az egyik leánysejtben nincs kromoszóma (aneuploidia), egy extra kromoszóma jelenléte (poliploidia), a a kromoszóma egy része (deléció) és átvitele egy másik kromoszómába (transzlokáció) . Ilyen folyamatok nagyon gyakran figyelhetők meg rosszindulatúan degenerált és transzformált sejtek szaporodása során. Ha ez a csírasejtek meiózisa során történik, az vagy a magzat halálához vezet az embrionális fejlődés korai szakaszában, vagy egy kromoszómabetegségben szenvedő szervezet megszületéséhez.
A tumornövekedés során a kontrollálatlan sejtszaporodás a sejtproliferációt szabályozó, onkogénnek nevezett gének mutációi miatt következik be. A jelenleg ismert több mint 70 onkogén közül a legtöbb a sejtnövekedés szabályozásának komponense, egy részük a génaktivitást szabályozó transzkripciós faktor, valamint a sejtosztódást és -növekedést gátló faktor. A szaporodó sejtek túlzott terjeszkedését (terjedését) korlátozó másik tényező a kromoszómák - telomerek - végeinek lerövidülése, amelyek a tisztán sztérikus kölcsönhatás következtében nem képesek teljesen replikálódni, ezért minden sejtosztódás után a telomerek lerövidülnek egy az alapok bizonyos része. Így egy felnőtt szervezet szaporodó sejtjei bizonyos számú osztódás után (általában 20-100, a szervezet típusától és életkorától függően) kimerítik a telomer hosszát, és a további kromoszóma replikáció leáll. Ez a jelenség nem fordul elő a spermatogén epitéliumban, az enterocitákban és az embrionális sejtekben a telomeráz enzim jelenléte miatt, amely minden osztódás után helyreállítja a telomerek hosszát. A legtöbb felnőtt szervezet sejtjében a telomeráz blokkolva van, de sajnos a daganatsejtekben aktiválódik.
A sejtmag és a citoplazma közötti kapcsolat, az anyagok mindkét irányú szállítása a nukleáris membrán pórusain keresztül történik, speciális energiafogyasztású szállítórendszerek részvételével. Így energia és képlékeny anyagok, szignálmolekulák (transzkripciós faktorok) kerülnek a magba. A fordított áramlás a citoplazmába juttatja az mRNS molekuláit és transzfer RNS-t (tRNS), a sejtben a fehérjeszintézishez szükséges riboszómákat. Ugyanez az anyagszállítási mód jellemző a vírusokra, különösen a HIV-re. Genetikai anyagukat a gazdasejt magjába juttatják, annak további beépülésével a gazda genomjába, és az újonnan képződött vírus RNS-t a citoplazmába juttatják az új vírusrészecskék további fehérjeszintéziséhez.
A szintézis folyamatainak megsértése
A fehérjeszintézis folyamatai az endoplazmatikus retikulum ciszternáiban mennek végbe, amelyek szorosan kapcsolódnak a magmembrán pórusaihoz, amelyeken keresztül a riboszómák, a tRNS és az mRNS bejutnak az endoplazmatikus retikulumba. Itt polipeptid láncok szintézisét végzik, amelyek később az agranuláris endoplazmatikus retikulumban és a lamelláris komplexumban (Golgi komplexben) nyerik el végső formájukat, ahol poszttranszlációs módosuláson és szénhidrát- és lipidmolekulákkal való asszociáción mennek keresztül. Az újonnan képződött fehérjemolekulák nem a szintézis helyén maradnak, hanem egy komplex szabályozott folyamat segítségével, amely ún. fehérje kinézis, aktívan átkerülnek a sejt azon izolált részébe, ahol ellátják tervezett funkciójukat. Ebben az esetben nagyon fontos lépés az átvitt molekula olyan megfelelő térbeli konfigurációvá strukturálása, amely képes ellátni benne rejlő funkcióját. Az ilyen strukturálás speciális enzimek segítségével, vagy speciális fehérjemolekulák - chaperonok - mátrixán történik, amelyek elősegítik, hogy az újonnan képződött vagy külső hatás hatására megváltozott fehérjemolekula a megfelelő háromdimenziós szerkezetet megszerezze. A sejtre gyakorolt káros hatás esetén, amikor fennáll a fehérjemolekulák szerkezetének megsértésének lehetősége (például testhőmérséklet-emelkedés, fertőző folyamat, mérgezés), a chaperonok koncentrációja a sejtben meredeken növekszik. Ezért az ilyen molekulákat is nevezik stresszfehérjék, vagy hősokk fehérjék. A fehérje molekula szerkezetének megsértése kémiailag inert konglomerátumok kialakulásához vezet, amelyek a sejtben vagy azon kívül rakódnak le, ha amiloidózis, Alzheimer-kór stb. hibás lesz. Ez a helyzet az úgynevezett prionbetegségeknél (juhsurlókór, tehén veszettség, kuru, Creutzfeldt-Jakob-kór emberben) fordul elő, amikor az idegsejt egyik membránfehérjéjének hibája inert tömegek későbbi felhalmozódását okozza a sejten belül. és létfontosságú tevékenységének megzavarása.
A sejtben a szintézis folyamatok megsértése annak különböző szakaszaiban fordulhat elő: RNS transzkripció a sejtmagban, polipeptidek transzlációja a riboszómákban, poszttranszlációs módosulás, a bézs színű molekula hipermetilációja és glikozilációja, fehérjék szállítása és eloszlása a sejtben, valamint eltávolításuk kifelé. Ebben az esetben megfigyelhető a riboszómák számának növekedése vagy csökkenése, a poliriboszómák lebomlása, a szemcsés endoplazmatikus retikulum ciszternáinak tágulása, a riboszómák elvesztése, vezikulák és vakuolák képződése. Tehát halvány gombagomba mérgezése esetén az RNS polimeráz enzim károsodik, ami megzavarja a transzkripciót. A diftéria toxin, amely inaktiválja az elongációs faktort, megzavarja a transzlációs folyamatokat, ami a szívizom károsodását okozza. Egyes specifikus fehérjemolekulák szintézisének megsértésének oka fertőző ágensek lehetnek. Például a herpeszvírusok gátolják az MHC antigénmolekulák szintézisét és expresszióját, ami lehetővé teszi számukra, hogy részben elkerüljék az immunkontrollt, a pestisbacilusok pedig gátolják az akut gyulladásközvetítők szintézisét. A szokatlan fehérjék megjelenése megállíthatja további lebomlását, és inert vagy akár mérgező anyagok felhalmozódásához vezethet. Ehhez bizonyos mértékig a bomlási folyamatok megzavarása is hozzájárulhat.
A bomlási folyamatok megsértése
A sejtben a fehérje szintézisével egyidejűleg annak bomlása folyamatosan megy végbe. Normál körülmények között ennek fontos szabályozó és formáló jelentősége van, például az enzimek inaktív formáinak, fehérjehormonoknak és a mitotikus ciklus fehérjéinek aktiválása során. A normál sejtnövekedés és -fejlődés finoman szabályozott egyensúlyt igényel a fehérjék és organellumok szintézise és lebontása között. A fehérjeszintézis folyamatában azonban a szintetizáló berendezés működésének hibái, a fehérjemolekula rendellenes szerkezete, kémiai és bakteriális szerek általi károsodása miatt folyamatosan nagyszámú hibás molekula képződik. Egyes becslések szerint részesedésük az összes szintetizált fehérje körülbelül egyharmada.
Az emlőssejteknek számos fő fehérje lebontási útvonalak: lizoszómális proteázokon (pentid-hidrolázokon), kalcium-dependens proteinázokon (endopeptidázokon) és a proteaszóma rendszeren keresztül. Ezen kívül léteznek speciális proteinázok is, például kaszpázok. A fő organellum, amelyben az eukarióta sejtekben az anyagok lebomlása megy végbe, a lizoszóma, amely számos hidrolitikus enzimet tartalmaz. A lizoszómák és fagolizoszómák endocitózisos folyamatai és különböző típusú autofágia következtében mind a hibás fehérjemolekulák, mind a teljes organellumok elpusztulnak: sérült mitokondriumok, plazmamembrán szakaszok, egyes extracelluláris fehérjék, szekréciós szemcsék tartalma.
A fehérjelebontás egyik fontos mechanizmusa a proteaszóma, egy összetett multikatalitikus proteináz szerkezet, amely a citoszolban, a sejtmagban, az endoplazmatikus retikulumban és a sejtmembránon lokalizálódik. Ez az enzimrendszer felelős a sérült fehérjék lebontásáért, valamint az egészséges fehérjékért, amelyeket el kell távolítani a normál sejtműködéshez. Ebben az esetben a megsemmisítendő fehérjéket előzetesen egy specifikus ubiquitin polipeptiddel kombinálják. Azonban a nem mindenütt jelenlévő fehérjék is részben elpusztulhatnak a proteaszómákban. A proteaszómákban lévő fehérjemolekulák rövid polipeptidekké történő lebomlása (feldolgozása), majd azok későbbi bemutatása az MHC I-es típusú molekulákkal együtt fontos láncszem a szervezet antigén homeosztázisának immunkontrolljának megvalósításában. A proteaszóma működésének gyengülése esetén a sérült és szükségtelen fehérjék felhalmozódása következik be, ami együtt jár a sejtek öregedésével. A ciklinfüggő fehérjék lebomlásának megsértése a sejtosztódás megsértéséhez, a szekréciós fehérjék lebomlásához vezet - cisztofibrózis kialakulásához. Ezzel szemben a proteaszóma működésének növekedése kíséri a szervezet kimerülését (AIDS, rák).
A fehérjelebontás genetikailag meghatározott megsértésével a szervezet nem életképes, és az embriogenezis korai szakaszában meghal. Ha a zsírok vagy szénhidrátok lebontása zavart okoz, akkor felhalmozódási betegségek (thesaurismosis) lépnek fel. Ugyanakkor a sejt belsejében túlzott mennyiségben halmozódnak fel bizonyos anyagok vagy azok hiányos bomlástermékei - lipidek, poliszacharidok -, ami jelentősen károsítja a sejt működését. Leggyakrabban máj epitheliocytákban (hepatocitákban), neuronokban, fibroblasztokban és makrofagocitákban figyelhető meg.
Az anyagok bomlási folyamatainak szerzett rendellenességei kóros folyamatok (például fehérje-, zsír-, szénhidrát- és pigment-disztrófia) eredményeként jelentkezhetnek, és szokatlan anyagok képződésével járhatnak együtt. A lizoszómális proteolízis rendszerének megsértése az alkalmazkodás csökkenéséhez vezet az éhezés vagy a megnövekedett terhelés során, bizonyos endokrin diszfunkciók előfordulásához - az inzulin, a tiroglobulin, a citokinek és receptoraik szintjének csökkenéséhez. A fehérjelebontás megsértése lassítja a sebgyógyulás ütemét, érelmeszesedés kialakulását idézi elő, és befolyásolja az immunválaszt. Hipoxia esetén az intracelluláris pH változása, sugárzási sérülés, amelyet a membránlipidek fokozott peroxidációja jellemez, valamint lizoszomotróp anyagok - bakteriális endotoxinok, mérgező gombák metabolitjai (sporofusarin), szilícium-oxid kristályok - hatása alatt a lizoszóma membrán stabilitása változások, aktivált lizoszómális enzimek szabadulnak fel a citoplazmába, ami a sejtszerkezetek pusztulását és halálát okozza.
1. fejezet
A SEJTÉLETTANI ALAPOK
I. Dudel
plazma membrán . Az állati sejteket a plazmamembrán korlátozza (1.1. ábra). Részletesebben fogunk foglalkozni szerkezetével, amely nagyon hasonló sok intracelluláris membrán szerkezetéhez. A membrán fő mátrixa abból áll lipidek főleg foszfatidilkolint. Ezek a lipidek egy hidrofil fejcsoportból állnak, amelyhez hosszú hidrofób szénhidrogénláncok kapcsolódnak. A vízben az ilyen lipidek spontán módon kétrétegű, 4-5 nm vastag filmet alkotnak, amelyben a hidrofil csoportok a vizes közeggel szemben állnak, a hidrofób szénhidrogénláncok pedig két sorban helyezkednek el, vízmentes lipidfázisot alkotva. A sejtmembránok ilyen típusú lipid kettős rétegek, és glikolipideket, koleszterint és foszfolipideket tartalmaznak (1.2. ábra). A glikolipidek hidrofil részét oligoszacharidok alkotják. A glikolipidek mindig a plazmamembrán külső felületén helyezkednek el, és a molekula oligoszacharid része a környezetbe merülő hajszálhoz hasonlóan orientált. A foszfolipidek között szétszórva közel azonos mennyiségű koleszterin molekula stabilizálja a membránt. A különböző lipidek eloszlása a membrán belső és külső rétegében nem azonos, sőt egyazon rétegen belül is vannak olyan területek, ahol bizonyos típusú lipidek koncentrálódnak. Ilyen egyenetlen eloszlás
Rizs. 1.1. Egy sejt sematikus rajza, amely a legfontosabb organellumokat mutatja
valószínűleg van némi, egyelőre homályos funkcionális jelentősége.
A membrán viszonylag inert lipidmátrixába merülő fő funkcionális elemek az mókusok(1.2. ábra). A fehérje tömege 25-75% a különböző membránokban, de mivel a fehérjemolekulák sokkal nagyobbak, mint a lipidmolekulák, 50 tömeg% 1 fehérjemolekula 50 lipidmolekula arányának felel meg. Egyes fehérjék áthatolnak a membránon annak külső felületéről a belső felületre, míg mások egy rétegben rögzülnek. A fehérjemolekulák általában úgy vannak orientálva, hogy hidrofób csoportjaik a lipidmembránba, a membrán felületén lévő poláris hidrofil csoportok pedig a vizes fázisba merüljenek. Számos külső felszíni membránfehérje glikoprotein; hidrofil szacharid csoportjaik az extracelluláris környezet felé néznek.
Az intracelluláris organellumok membránrendszerei .
A sejttérfogat körülbelül felét a citoszolból membránokkal izolált organellumok foglalják el. Az intracelluláris organellumok membránjainak teljes felülete legalább 10-szerese a plazmamembrán felületének. A legszélesebb körben használt membránrendszer az endoplazmatikus retikulum, hálózatot képvisel

Rizs. 1.2.A plazmamembrán sematikus ábrázolása. A fehérjék a foszfolipid kettős rétegbe vannak beágyazva, egyesek átfogják a kettős réteget, míg mások csak a külső vagy belső réteghez vannak rögzítve.
erősen csavarodott tubulusok vagy zsákszerű hosszúkás szerkezetek; az endoplazmatikus retikulum nagy területeit riboszómák tarkítják; az ilyen retikulumot szemcsésnek vagy érdesnek nevezzük (1.1. ábra). golgi készülék szintén membránhoz kötött lamellákból áll, amelyekről hólyagok, vagy hólyagok törnek le (1.1. ábra). Lizoszómák és peroxiszómák kis speciális hólyagok. Mindezekben a változatos organellumokban a membrán és az általa lefedett tér meghatározott enzimkészleteket tartalmaz; az organellumok belsejében speciális anyagcseretermékek halmozódnak fel, amelyek az organellumok különféle funkcióinak ellátására szolgálnak.
MagÉs mitokondriumok abban különböznek, hogy ezen organellumok mindegyikét két membrán veszi körül. A sejtmag felelős az anyagcsere kinetikai szabályozásáért; az összehajtogatott belső mitokondriális membrán az oxidatív anyagcsere helyszíne; itt a piruvát vagy a zsírsavak oxidációja következtében egy nagy energiájú vegyület adenozin-trifoszfát (ATP, vagy ATP) szintetizálódik.
citoszkeleton . Az organellumokat körülvevő citoplazma semmiképpen sem tekinthető amorfnak; citoszkeleton hálózata hatja át. A citoszkeleton mikrotubulusokból, aktinszálakból és közbenső filamentumokból áll (1.1. ábra). mikrotubulusok külső átmérőjük körülbelül 25 nm; normál polimerhez hasonlóan tubulin fehérje molekulák összeállítása eredményeként jönnek létre. aktin filamentumok - a membránközeli rétegben és az egész sejtben elhelyezkedő kontraktilis rostok - főként a mozgással kapcsolatos folyamatokban vesznek részt. Köztes szálak különböző típusú sejtekben különböző kémiai összetételű blokkokból áll; a citoszkeleton fent említett másik két eleme között sokféle kapcsolatot alkotnak. A citoszkeletonhoz az organellumok és a plazmamembrán is társul, amely nemcsak a sejt alakját és a benne lévő organellumok helyzetét tartja fenn, hanem meghatározza a sejt alakjának változását és mobilitását is.
Citoszol . A sejttérfogat körülbelül felét a citoszol foglalja el. Mivel körülbelül 20% (tömeg) fehérjét tartalmaz, inkább gél, mint vizes oldat. Kis molekulák, beleértve a szerves és szervetlen molekulákat ionok, feloldjuk a vizes fázisban. A sejt és a környezet (extracelluláris tér) között ioncsere zajlik; ezekről a cserefolyamatokról a következő részben lesz szó. Az ionok koncentrációját az extracelluláris térben jelentős pontossággal állandó szinten tartják; az egyes ionok intracelluláris koncentrációja is rendelkezik egy sajátos szinttel, amely eltér a sejten kívülitől (1.1. táblázat). Az extracelluláris környezetben a leggyakoribb kation az Na+ a sejtben koncentrációja több mint 10-szer alacsonyabb. Ezzel szemben a sejten belül a legmagasabb a K + koncentrációja, a sejten kívül pedig több mint egy nagyságrenddel alacsonyabb. A legnagyobb gradiens az extracelluláris és intracelluláris koncentráció között a Ca 2+ esetében van, amelynek a sejten belüli szabad ionjainak koncentrációja legalább 10 000-szer kisebb, mint azon kívül. Nem minden ion oldódik fel a citoszolban, egy részük fehérjéken adszorbeálódik vagy organellumokban rakódik le. Például a Ca 2+ esetében a kötött ionok sokkal többen vannak, mint a szabadok. A citoszolfehérjék többsége enzim, amelyek részvételével számos köztes anyagcsere-folyamat megy végbe: glikolízis és glükoneogenezis, aminosavak szintézise vagy elpusztítása, fehérjeszintézis riboszómákon (1.1. ábra). A citoszol lipidcseppeket és glikogén granulátumokat is tartalmaz, amelyek fontos molekulák tartalékaiként szolgálnak.
1.1. táblázat.Az ionok intra- és extracelluláris koncentrációi homoioterm állatok izomsejtjeiben. A – – „nagy molekulatömegű sejtanionok”
|
Intracelluláris koncentráció |
Extracelluláris koncentráció |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
l-el- |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO 3 - |
8 mmol/l |
HCO 3 - |
27 mmol/l |
|
A- |
155 mmol/l |
Egyéb kationok |
5 mmol/l |
|
Nyugalmi potenciál -90 mV |
|||
1.2. Anyagcsere a sejt és a környezet között
Röviden ismertettük a sejt szerkezetét, hogy ezzel a leírással áttekinthessük a sejtfiziológia alapjait. Egy sejt semmi esetre sem tekinthető statikus képződménynek, mivel állandó anyagcsere zajlik a különböző intracelluláris kompartmentek, valamint a kompartmentek és a környezet között. A sejt szerkezetei dinamikus egyensúlyban vannak, a sejtek egymással és a külső környezettel való kölcsönhatása elengedhetetlen feltétele a működő szervezet életének fenntartásához. Ebben a fejezetben megvizsgáljuk az ilyen csere alapvető mechanizmusait. A későbbi fejezetekben ezeket a mechanizmusokat az idegsejttel és annak funkcióival összefüggésben tárgyaljuk;
ugyanakkor ugyanazok a mechanizmusok állnak az összes többi szerv működésének hátterében.
Diffúzió.Az anyag mozgatásának legegyszerűbb folyamata a diffúzió. Az oldatokban (vagy gázokban) az atomok és molekulák szabadon mozognak, a koncentrációkülönbséget diffúzió egyensúlyozza ki. Vegyünk két folyadékkal vagy gázzal töltött térfogatot (1.3. ábra), amelyekben az anyagok koncentrációja van c1 és c2 és A felületű és vastagságú réteg választja el d. Az m anyag áramlása t időben leírta Fick első diffúziós törvénye:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
ahol D a diffúziós együttható, amely egy adott anyagra, oldószerre és hőmérsékletre állandó. Általánosabb formában a koncentrációkülönbségre dc távolságra dx
dm/dt= –D A dc/dx,(2)
az A szakaszon keresztüli áramlás arányos a koncentrációgradienssel dc/dx . A mínusz előjel azért jelenik meg az egyenletben, mert a koncentráció x irányú változása negatív.
A diffúzió a legfontosabb folyamat, amelynek során a vizes oldatokban lévő molekulák többsége rövid távolságra mozog. Ez vonatkozik a sejtben való mozgásukra is, amennyiben a diffúziót nem akadályozzák a membránok. Számos anyag szabadon átdiffundálhat a lipidmembránokon, különösen a víz és az oldott gázok, például az O 2 és a CO 2 . zsírban oldódó

Rizs. 1.3.A diffúzió mennyiségi sémája. A két teret vastagságréteg választja eldés terület A. C; - magas részecskekoncentráció a térfogat bal oldalán, C:, - alacsony részecskekoncentráció a jobb oldalon részek, rózsaszín felület a koncentráció gradiens a diffúziós rétegben. Diffúziós fluxus dm/dt – lásd (1) egyenlet
az anyagok a membránokon is jól diffundálnak; ez vonatkozik a viszonylag kis poláris molekulákra is, mint az etanol és a karbamid, míg a cukrok nehezen jutnak át a lipidrétegen. Ugyanakkor a lipidrétegek gyakorlatilag áthatolhatatlanok a töltött molekulák számára, beleértve még a szervetlen ionokat is. Nem elektrolitok esetében az (1) diffúziós egyenletet általában úgy alakítják át, hogy a membrán és a diffundáló anyag jellemzőit egyesítik. paraméter-permeabilitás (P):
dm/dt=P AD c.(3)
ábrán. 1,4 összehasonlítva a lipidmembrán permeabilitása (P) különböző molekulák számára.
Diffúzió a membrán pórusain keresztül . A plazmamembrán (és más sejtmembránok) nemcsak a lipidrétegen átdiffundáló anyagok, hanem számos ion, cukor, aminosav és nukleotid számára is átjárható. Ezek az anyagok átjutnak a membránon az általa kialakított pórusokon keresztül transzport fehérjék, membránba ágyazva. Az ilyen fehérjéken belül egy 1 nm-nél kisebb átmérőjű vízzel teli csatorna található, amelyen keresztül a kis molekulák diffundálhatnak. Koncentrációgradiens mentén mozognak, és ha töltést hordoznak, akkor a csatornákon való mozgásukat a membránpotenciál is szabályozza. A membráncsatornák viszonylag szelektívek

Rizs. 1.4.Mesterséges lipid kettősrétegek permeabilitása különböző anyagokhoz
A rajtuk áthaladó molekulák típusával kapcsolatban vannak például kálium-, nátrium- és kalciumcsatornák, amelyek mindegyike szinte minden ion számára áthatolhatatlan, kivéve egy bizonyos iont. Ilyen szelektivitás a csatorna falában lévő kötőhelyek töltése vagy szerkezete miatt, ami megkönnyíti egy adott molekula szállítását és megakadályozza más anyagok behatolását a csatornán keresztül (2. ábra). 1,5, A) .
A viselkedés mögött membrán ioncsatornák Könnyen megfigyelhető, hiszen az ionok mozgása során fellépő áramerősség mérhető, és akár egyetlen csatornára is. Megmutatták, hogy a csatornák spontán és nagy frekvenciával változtatják állapotukat nyitottról zártra. A káliumcsatornát körülbelül 2 pA (2 10 -12 A) amplitúdójú és több milliszekundumos időtartamú áramimpulzusok jellemzik (lásd 2.12. ábra, 37. o.) [3]. Ebben az időszakban több tízezer ion halad át rajta. A fehérjék egyik konformációból a másikba való átmenetét röntgendiffrakcióval, Mössbauer-spektroszkópiával és mágneses magrezonanciával (NMR) vizsgálják. A fehérjék tehát rendkívül dinamikus, mobil struktúrák, és a fehérjéken áthaladó csatorna nem csupán egy merev, vízzel teli cső (1.5A ábra), hanem gyorsan mozgó molekulacsoportok és töltések labirintusa. Ez a csatorna dinamikus válasza tükröződik a csatorna energiaprofilja,ábrán látható. 1.5, B. Itt az abszcissza a C 0 ionkoncentrációjú és 0 potenciálú külső oldattól a C 1 koncentrációjú és E potenciálú belső oldatig vezető csatorna hosszát mutatja. Az y tengely

Rizs. 1.5.A. A plazmamembrán lipid kettős rétegébe ágyazott káliumcsatornát alkotó fehérje vázlata. A csatorna "falán" négy negatív töltés van rögzítve. B. ábrán látható csatorna sematikus energiaprofilja. A. Az y tengelyen a csatorna áthaladásához szükséges kinetikus energia értékeit ábrázoljuk; az abszcissza tengely mentén a membrán belső és külső felülete közötti távolságok. Az energiaminimumok a pozitív töltésű, fix negatív töltésű ionok kötőhelyeinek felelnek meg a csatorna falában. Az energiamaximumok a csatornában lévő diffúziós akadályoknak felelnek meg. Feltételezzük, hogy a csatornafehérje konformációja spontán oszcillál; az energiaprofilok lehetőségeit folyamatos és szaggatott vonalak jelzik; ezek az oszcillációk nagymértékben megkönnyítik az ionok megkötését az energiagát leküzdésekor (de változásokkal)
az ion energiaszintjeit a csatornakötő helyeken mutatjuk be; a grafikon csúcsa azt a permeabilitási gátat jelöli, amelyet az ionenergiának le kell győznie ahhoz, hogy áthatoljon a csatornán, a gráf „bemerülése” pedig egy viszonylag stabil állapotot (kötést). Az energiacsúcs-elzáródás ellenére egy ion áthatolhat a csatornán, ha az energiaprofil spontán ciklusokat vált ki; így az ion hirtelen megjelenhet az energiacsúcs "túloldalán", és tovább tud mozogni a sejtbe. Az ion töltésétől, méretétől, hidratáltsági fokától, valamint a csatornafalak szerkezetéhez való kötődési képességétől függően a csatorna energiaprofilja a különböző ionoknál eltérő, ami magyarázhatja az egyes csatornatípusok szelektivitását.
Ionok diffúziós egyensúlya . A különböző ionok diffúziója a membráncsatornákon az extracelluláris és intracelluláris környezet közötti koncentrációbeli különbségek kiküszöböléséhez vezet. Amint azonban a táblázatból látható. 1.1, ezek a különbségek továbbra is fennállnak, tehát biztosan van néhány egyensúlyi diffúzió és más transzportfolyamatok között a membránon keresztül. A következő két rész az egyensúly megteremtésének módjaival foglalkozik. Ionok esetében a diffúziós egyensúlyt töltésük befolyásolja. A töltetlen molekulák diffúzióját a koncentrációkülönbség biztosítja dc , és amikor a koncentrációk kiegyenlítődnek, a tényleges szállítás leáll. A töltött részecskékre ezenkívül az elektromos tér is hatással van. Például, amikor egy káliumion kilép a sejtből a koncentrációgradiense mentén, egy pozitív töltést hordoz. Így az intracelluláris környezet negatívabb töltésűvé válik, ami potenciálkülönbséget eredményez a membránon keresztül. Az intracelluláris negatív töltés megakadályozza, hogy új káliumionok távozzanak a sejtből, és azok az ionok, amelyek ennek ellenére elhagyják a sejtet, tovább növelik a membrán töltését. A káliumionok áramlása leáll, amikor az elektromos tér hatása kompenzálja a koncentrációkülönbség miatti diffúziós nyomást. Az ionok továbbra is áthaladnak a membránon, de mindkét irányban egyenlő mennyiségben. Ezért a membránon lévő ionkoncentrációk adott különbségére létezik egyensúlyi potenciál E ion amelynél leáll az ionok áramlása a membránon keresztül. Az egyensúlyi potenciál könnyen meghatározható a segítségével Nernst egyenletek:
Eion= RT/ zF* lnKi/ C be(4)
ahol R a gázállandó, T az abszolút hőmérséklet, z az ion vegyértéke (anionokra negatív) Ki az extracelluláris ionkoncentráció, C be az ion intracelluláris koncentrációja, F Faraday szám. Ha az egyenletbe állandókat helyettesítünk, akkor testhőmérsékleten (T = 310 K) a káliumionok egyensúlyi potenciálja E K egyenlő:
Ek= –61 mB log /(5)
Ha [ K + out ]/[ K + in ] = 39, a táblázat szerint. akkor 1.1
Ek= -61 m B log 39= -97 mV.
Valójában azt találták, hogy minden sejt rendelkezik Membránpotenciál; emlős izomsejtekben szintje körülbelül -90 mV. A körülményektől és az ionok relatív koncentrációjától függően a sejtek membránpotenciálja -40 és -120 mV között lehet. A fenti példa cellájához (1.1. táblázat) nyugalmi potenciál, körülbelül -90 mV, azt jelzi, hogy a membráncsatornákon áthaladó káliumionok fluxusa megközelítőleg egyensúlyban van. Ez nem meglepő, hiszen a nyugalmi membránban a káliumcsatornák nyitott állapota a legvalószínűbb; a membrán a leginkább áteresztő a káliumionok számára. A membránpotenciált azonban más ionok fluxusai is meghatározzák.
A (3) egyenletben számszerűsíthető, hogy a töltetlen részecskék milyen könnyen átdiffundálhatnak a membránon. A töltött részecskék áteresztőképessége egy kicsit bonyolultabb egyenlettel írjuk le:
P= m RT/ dF(6)
Ahol maz ion mobilitása a membránban, d - membrán vastagsága, egy R, T és F ismert termodinamikai állandók. Az így meghatározott különböző ionok permeabilitási értékei felhasználhatók a membránpotenciál kiszámításához Em amikor a kálium-, nátrium- és kloridionok egyidejűleg haladnak át a membránon (P permeabilitással K, P Na és P Cl illetőleg). Feltételezzük, hogy a potenciál egyenletesen csökken a membránban, így a térerősség állandó. Ebben az esetben ez vonatkozik Goldman-egyenlet, vagy állandó mezőegyenlet :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl ) (7)
A legtöbb sejtmembránhoz P K körülbelül 30-szor magasabb, mint az R Na (lásd még az 1.3. pontot). Relatív érték PCl nagymértékben változik; sok membránhoz PCl kicsi R-hez képest K azonban mások számára (pl. a vázizomzatban) PCl , sokkal magasabb, mint az R K.
Aktív szállítás, nátriumpumpa . Az előző rész az ionok passzív diffúzióját és az ebből eredő membránpotenciálját írja le adott intra- és extracelluláris ionkoncentrációk mellett. Ennek a folyamatnak az eredményeként azonban a sejten belüli ionok koncentrációja nem stabilizálódik automatikusan, mivel a membrán
potenciál valamivel elektronegatívabb, mint E K , és sokkal több E-hez képest Na (kb. +60 mV). A diffúzió miatt az intracelluláris ionok, legalább a kálium és a nátrium koncentrációjának ki kell egyenlítenie az extracellulárisakkal. Az iongradiens stabilitása az aktív transzport révén érhető el: a membránfehérjék az elektromos és (vagy) koncentrációgradiensek ellenében szállítják az ionokat a membránon keresztül, ehhez metabolikus energiát fogyasztanak. Az aktív közlekedés legfontosabb folyamata a munka Na/K - egy szivattyú, amely szinte minden cellában létezik;
a szivattyú nátriumionokat pumpál ki a cellából, miközben káliumionokat pumpál a sejtbe. Ez biztosítja a nátriumionok alacsony intracelluláris koncentrációját és a magas káliumkoncentrációt (1.1. táblázat). A nátriumionok koncentráció-gradiense a membránon az információ elektromos impulzusok formájában történő továbbításához (lásd 2.2. fejezet), valamint egyéb aktív transzportmechanizmusok fenntartásához és a sejttérfogat szabályozásához (lásd alább) specifikus funkciókat lát el. Ezért nem meglepő, hogy a sejt által fogyasztott energia több mint 1/3-át a Na / K szivattyúra fordítják, és a legaktívabb sejtekben az energia akár 70% -át is a működésére fordítják.
A Na/K transzport fehérje egy ATP-áz. A membrán belső felületén az ATP-t ADP-re és foszfátra hasítja (1.6. ábra). Egy ATP-molekula energiája három nátriumion és egyidejűleg két káliumion szállítására szolgál a sejtből, azaz összesen egy pozitív töltés távozik a sejtből egy ciklus alatt. Így a Na/K szivattyú az elektrogén(elektromos áramot hoz létre a membránon keresztül), ami a membránpotenciál elektronegativitásának körülbelül 10 mV-os növekedéséhez vezet. A transzportfehérje ezt a műveletet nagy sebességgel hajtja végre: 150-600 nátriumion másodpercenként. A transzport fehérje aminosavsorrendje ismert, de ennek a komplex cseretranszportnak a mechanizmusa még nem tisztázott. Ezt a folyamatot a nátrium- vagy káliumionok fehérjék általi átvitelének energiaprofilja segítségével írjuk le (1.5.5. ábra). Ezeknek a profiloknak a transzportfehérje konformációjának állandó változásával (energiát igénylő folyamat) járó változás természetéből adódóan a kicserélődés sztöchiometriája megítélhető: két káliumion három nátriumionra cserélődik.
Na/K-szivattyú, mint szigetelt Na+ /K + -függő membrán-ATPáz, amelyet specifikusan az ouabain (sztrofantin) szívglikozid gátolja. Mivel a Na/K szivattyú működése többlépcsős kémiai reakció, ezért, mint minden kémiai reakció, nagymértékben hőmérsékletfüggő, ami

Rizs. 1.6.A Na/K-pumpa-ATPáz (a plazmamembrán lipid kettősrétegébe merülve) sémája, amely egy ciklus alatt három Na + iont vesz ki a sejtből a potenciál- és koncentráció gradiensek ellenében és két K iont visz be a sejtbe + . A folyamat során egy ATP-molekula ADP-re és foszfátra hasad. Az ábrán az ATPáz egy nagy (funkcionális) és egy kis alegységből álló dimerként látható; a membránban tetramerként létezik, amelyet két nagy és két kis alegység alkot
ábrán látható. 1.7. Itt az izomsejtekből származó nátriumionok áramlása látható az idő függvényében; ez gyakorlatilag megegyezik a Na/K szivattyú működése által közvetített nátriumion-áramlással, mert a nátriumionok passzív áramlása a koncentrációval és a potenciálgradiensekkel szemben rendkívül kicsi. Ha a készítményt kb. 18°C-ra lehűtjük, akkor a sejtből a nátriumionok áramlása gyorsan 15-szörösére csökken, és melegítés után azonnal visszaáll az eredeti szintre. A sejtből a nátriumionok áramlásának ilyen csökkenése többszöröse annak, ami a diffúziós folyamat vagy egy egyszerű kémiai reakció hőmérsékletfüggésének felelne meg. Hasonló hatás figyelhető meg, amikor a metabolikus energia kimerül a dinitrofenol (DNP) mérgezés következtében (1.7.5. ábra). Ezért a nátriumionok áramlását a sejtből egy energiafüggő reakció - egy aktív szivattyú - biztosítja. A szivattyú másik jellemzője a jelentős hőmérséklet- és energiafüggéssel együtt a telítési szint jelenléte (mint minden más kémiai reakció esetében); ez azt jelenti, hogy a szállított ionok koncentrációjának növekedésével a szivattyú fordulatszáma nem növekedhet korlátlanul (1.8. ábra). Ezzel szemben a passzívan diffundáló anyag áramlása a diffúzió törvényének megfelelően a koncentrációk különbségével arányosan nő (1. és 2. egyenlet).

Rizs. 1.7. A, B. Na aktív közlekedés + . Y tengely: radioaktív 24 Na + áramlása a sejtből (imp./perc). abszcissza tengely: a kísérlet kezdete óta eltelt idő. A. A cellát 18,3 °C-ról 0,5 °C-ra hűtjük; folyam Na+ Ebben az időszakban a sejtből való kilépés gátolt. B. A Na + sejtből való kiáramlásának elnyomása dinitrofenollal (DNF) 0,2 mmol/l koncentrációban (módosítva)
A Na / K pumpán kívül a plazmamembrán még legalább egy pumpát tartalmaz - kalcium; ez a pumpa kalciumionokat (Ca 2+) pumpál ki a sejtből, és részt vesz azok intracelluláris koncentrációjának rendkívül alacsony szinten tartásában (1.1. táblázat). A kalciumpumpa igen nagy sűrűséggel van jelen az izomsejtek szarkoplazmatikus retikulumában, amelyek az ATP molekulák lebontása következtében kalciumionokat halmoznak fel (lásd 4. fejezet).
A Na/K pumpa hatása a membránpotenciálra és a sejttérfogatra . ábrán. Az 1.9 a membránáram különböző összetevőit mutatja, és az ionok intracelluláris koncentrációit mutatja, amelyek

Rizs. 1.8.A molekulák szállítási sebessége és koncentrációjuk (a csatornába való belépés vagy a pumpa megkötésének pontján) aránya a csatornán keresztüli diffúzió vagy a szivattyúzás során. Ez utóbbi nagy koncentrációban telítődik (maximális sebesség, Vmax ) érték az abszcisszán, amely a szivattyú maximális fordulatszámának felének felel meg ( Vmax /2), az egyensúlyi koncentráció NAK NEK m

Rizs. 1.9.A Na+ koncentrációkat bemutató diagram , K+ és Cl- sejten belül és kívül, valamint ezen ionok sejtmembránon való áthatolási útvonalai (specifikus ioncsatornákon vagy a Na/K pumpa segítségével. Adott koncentráció gradiensek mellett az egyensúlyi potenciálok E Na, E K és E C l - megegyeznek a jelzettekkel, a membránpotenciál Em = – 90 mV
biztosítják létezésüket. A káliumcsatornákon keresztül a káliumionok kifelé irányuló árama figyelhető meg, mivel a membránpotenciál valamivel elektropozitívabb, mint a káliumionok egyensúlyi potenciálja. A nátriumcsatornák teljes vezetőképessége jóval alacsonyabb, mint a káliumcsatornáké; a nátriumcsatornák sokkal ritkábban nyílnak meg, mint a nyugalmi helyzetben lévő káliumcsatornák; ugyanakkor körülbelül ugyanannyi nátriumion lép be a sejtbe, mint amennyi a káliumion elhagyja, mivel a nátriumionok sejtbe történő diffúziójához nagy koncentráció és potenciálgradiens szükséges. A Na/K pumpa ideális kompenzációt biztosít a passzív diffúziós áramokhoz, mivel a nátriumionokat szállítja ki a sejtből és a káliumionokat abba. Így a szivattyú elektrogén a cellába be- és kikerülő töltések számának különbsége miatt, ami normál működési sebesség mellett körülbelül 10 membránpotenciált hoz létre. Az mV elektronegatívabb, mintha csak passzív ionáramok hoznák létre (lásd a 7. egyenletet). Ennek eredményeként a membránpotenciál megközelíti a kálium egyensúlyi potenciálját, ami csökkenti a káliumionok szivárgását. Na tevékenység A /K-pumpát a nátriumionok intracelluláris koncentrációja szabályozza. A pumpa sebessége a sejtből eltávolítandó nátriumionok koncentrációjának csökkenésével lelassul (1.8. ábra), így a pumpa működése és a nátriumionok sejtbe áramlása kiegyenlíti egymást, fenntartva az intracelluláris koncentrációt. nátriumionok mennyisége körülbelül 10 mmol / l.
A pumpáló és a passzív membránáramok közötti egyensúly fenntartásához sokkal több Na/K-pumpa molekulára van szükség, mint amennyi csatornafehérje a kálium- és nátriumionokhoz. Ha a csatorna nyitva van, néhány ezredmásodperc alatt több tízezer ion halad át rajta (lásd fent), és mivel a csatorna általában másodpercenként többször nyílik, ezalatt összesen több mint 10 5 ion halad át rajta. Egyetlen pumpafehérje több száz nátriumiont mozgat meg másodpercenként, így a plazmamembránnak körülbelül 1000-szer több pumpa molekulát kell tartalmaznia, mint a csatornamolekuláknak. A nyugalmi csatornaáramok mérése átlagosan egy kálium és egy nátrium nyitott csatornát mutatott 1 µm 2 membránonként; ebből az következik, hogy ugyanabban a térben körülbelül 1000 Na/K pumpa molekulának kell lennie, azaz. a köztük lévő távolság átlagosan 34 nm; a pumpáló fehérje, mint csatornafehérje átmérője 8-10 nm. Így a membrán kellően sűrűn telített pumpáló molekulákkal
Az a tény, hogy a nátriumionok beáramlását a sejtbe, és a káliumionok sejtből kiáramlását a szivattyú működése kompenzálja, egy másik következménnyel jár, ami az ozmotikus nyomás stabil fenntartása, ill. állandó hangerő. A sejt belsejében nagy koncentrációban találhatók nagy anionok, főleg fehérjék (A - az 1.1. táblázatban), amelyek nem képesek áthatolni a membránon (vagy nagyon lassan hatolnak be), ezért a sejt belsejében rögzített komponensek. Ezen anionok töltésének kiegyensúlyozásához azonos számú kationra van szükség. A Na/K pumpa hatására ezek a kationok főként káliumionok. Az ionok intracelluláris koncentrációjának szignifikáns növekedése csak az anionok koncentrációjának növekedése esetén következhet be a C1 - a koncentráció gradiens sejtbe áramlása miatti - növekedése mellett (1.1. táblázat), de a membránpotenciál ezt ellensúlyozza. Bejövő áram Cl- csak a kloridionok egyensúlyi potenciáljának eléréséig figyelhető meg; ez akkor figyelhető meg, ha a kloridion-gradiens majdnem ellentétes a kálium-ion-gradienssel, mivel a kloridionok negatív töltésűek (4. egyenlet). Így a kloridionok alacsony intracelluláris koncentrációja jön létre, ami megfelel a káliumionok alacsony extracelluláris koncentrációjának. Az eredmény a sejtben lévő ionok teljes számának korlátozása. Ha a Na/K pumpa blokkolásakor a membránpotenciál csökken, például anoxia során, akkor a kloridionok egyensúlyi potenciálja csökken, és ennek megfelelően nő a kloridionok intracelluláris koncentrációja. A töltések egyensúlyának helyreállításával a káliumionok is bejutnak a sejtbe; nő az ionok összkoncentrációja a sejtben, ami növeli az ozmotikus nyomást; ez arra kényszeríti a vizet, hogy belépjen a sejtbe. A sejt megduzzad. Ez a duzzanat látható in vivo energiahiányos körülmények között.
koncentráció gradiens Na + mint a membrántranszport hajtóereje . A Na/K-pumpa jelentősége a sejt számára nem korlátozódik a normál K+ és Na+ gradiensek stabilizálására a membránon keresztül. A membrán gradiensben tárolt energia Na+ , gyakran használják más anyagok membrántranszportjának biztosítására. Például a 2. ábrán. 1.10 mutatja a "szimportot" Na+ és a cukormolekulák a sejtbe. A membrán transzport fehérje a cukormolekulát a koncentrációgradiens ellenében is beviszi a sejtbe, míg a Na + a koncentráció és a potenciál gradiens mentén mozog, energiát adva cukrok szállítása. A cukrok ilyen szállítása teljes mértékben a magas gradiens meglététől függ Na+ ; ha az intracelluláris koncentráció Na+ jelentősen megnő, akkor a cukrok szállítása leáll. Különféle c Akharov, különböző szimportrendszerek léteznek. Az aminosavak szállítása a szállításhoz hasonló ketrecben c ábrán látható akharov. 1,10; a gradiens is biztosítja Na+ , legalább öt különböző szimportrendszer létezik, amelyek mindegyike rokon aminosavak egy-egy csoportjára specializálódott.
A szimportrendszereken kívül vannak még "antiport". Egyikük például egy ciklus alatt egy kalciumiont visz ki a sejtből három bejövő nátriumionért cserébe (1.10. ábra). Energia a szállítás Ca 2+ három nátriumion bejutása következtében jön létre a koncentráció és a potenciálgradiens mentén. Ez az energia elegendő (nyugalmi potenciálon) a magas kalciumion-gradiens fenntartásához (kevesebb, mint 10-7 mol/l-ről a sejten belül körülbelül 2 mmol/l-re a sejten kívül).
Endo- és exocitózis . Bizonyos anyagokra, amelyek bejutnak a sejtbe, vagy el kell távolítani

Rizs. 1.10.A membrán lipid kettős rétegébe ágyazott fehérjék közvetítik a glükóz és a Na + sejtbe történő bejutását, valamint Ca2+/Na+ -antiport, amelyben a hajtóerő a Na + gradiense a sejtmembránon
belőle nincsenek szállítási csatornák; ilyen anyagok például a fehérjék és a koleszterin. A plazmamembránon keresztül bejuthatnak hólyagok vagy buborékok, endo- és exocitózissal. ábrán. Az 1.11 bemutatja e folyamatok főbb mechanizmusait. Az exocitózis során bizonyos organellumok (lásd alább) hólyagokat képeznek, amelyek tele vannak olyan anyaggal, amelyet el kell távolítani a sejtből, például hormonokkal vagy extracelluláris enzimekkel. Amikor az ilyen vezikulák elérik a plazmamembránt, lipidmembránjuk összeolvad vele, így lehetővé teszi a tartalom kijutását a külső környezetbe. Ezzel ellentétes folyamatban, az endocitózisban, a plazmamembrán behatol, gödröt képez, amely azután mélyül és bezárul, és extracelluláris folyadékkal és néhány makromolekulával feltöltött intracelluláris vezikulát képez. A membránösszeolvadás és a vezikula záródása érdekében a citoszkeleton összehúzódó elemei magukkal a membránokkal együtt hatnak (lásd alább). Az endocitózis nem mindig jelenti az extracelluláris közegnek a sejtbe való befogását. A sejtmembrán gyakran speciális csoportokba rendezve tartalmaz makromolekulák, például inzulin vagy antigének specifikus receptorait. Miután ezek a makromolekulák a receptoraikhoz kötődnek, a receptort körülvevő membrán régióban endocitózis lép fel, és a makromolekula szelektíven bejut a sejtbe (1.12. ábra, B).
Az endo- és exocitózis folyamatosan fordul elő a sejtekben. Jelentős a keringő membránanyag mennyisége; 1 órán belül a makrofág vezikulák formájában abszorbeálja a citoplazma membrán felületének kétszeresét. A legtöbb sejtben a membránanyag forgalom nem olyan intenzív, de ennek ellenére jelentősnek kell lennie.

Rizs. 1.11.exocitózis és endocitózis. Fel: az intracelluláris vezikula összeolvad a plazmamembrán lipid kettős rétegével és az extracelluláris térbe nyílik. Ezt a folyamatot exocitózisnak nevezik. Az alján: a plazmamembrán kis területen invaginál, és egy extracelluláris anyaggal teli vezikulát fűz le. Ezt a folyamatot endocitózisnak nevezik.
1.3. Anyagok szállítása a sejten belül
Az endo- és exocitózis nemcsak az anyagok sejtmembránon keresztül történő szállításának folyamata, hanem a membráncsere folyamatai is - a sejt szerkezeti összetevői. Ebben a részben a sejtben és sejtszervecskéiben végbemenő egyéb hasonló transzportfolyamatokról van szó.

Rizs. 1.12. A-B. A folyamatok sémája, beleértve az exo- és endocitózist. A. A granuláris endoplazmatikus retikulumban szintetizált fehérje a Golgi-készüléken keresztül a plazmamembránba kerül, ahol exocitózissal kiválasztódik. B. Az LDL (alacsony sűrűségű lipoprotein) részecskékhez kötött koleszterin a plazmamembránhoz tapad, endocitikus vezikula képződését idézi elő a membrán ezen régiójában, és a lizoszómákba kerül, ahol felszabadul. BAN BEN. Az endocitózis során befogott extracelluláris anyag (az ábrán jobb oldalon), vezikulákban vagy vezikulákban szállítják a sejten keresztül, és exocitózissal szabadulnak fel (az ábrán bal)
Diffúzió . Természetesen a citoszolban a koncentrációkülönbség diffúzióval megszűnik; ugyanez igaz az organellumokban lévő folyadékokra is. Az oldott fehérje magas koncentrációja miatt a diffúzió itt sokkal lassabb, mint a vízben. A lipidmembránok – a sejt körül és az organellumokon belül – kétdimenziós folyadékok, amelyekben diffúzió megy végbe. A membrán kettős rétegében lévő lipidek saját rétegükön belül diffundálnak, ritkán mozognak egyikből a másikba. A bennük elmerült fehérjék is meglehetősen mozgékonyak; a membránra merőleges tengely körül forognak, vagy oldalirányban diffundálnak nagyon eltérő diffúziós állandókkal, 2-10 000-szer lassabban, mint a foszfolipidek. Tehát, ha egyes fehérjék szabadon mozognak a lipidrétegben és ugyanolyan sebességgel, mint maguk a lipidmolekulák, akkor mások lehorgonyoznak, pl. elég erősen kapcsolódik a citoszkeletonhoz. A membránban specifikus fehérjék „permanens” aggregátumai vannak, például az idegsejtek pre- és posztszinaptikus struktúrái. A szabadon mozgó fehérjéket fluoreszcens festékekhez kötve lehet kimutatni, amelyeket a membrán egy kis részének rövid villanásokkal történő rövid megvilágításával késztetnek ragyogásra. Az ilyen kísérletek azt mutatják, hogy kevesebb, mint 1 perc alatt a festékhez kötött fehérjék egyenletesen oszlanak el a membránon 10 μm távolságig.
Aktív transzport az organellummembránokban .
A plazmamembrán működésében létfontosságú szerepet játszó aktív transzport folyamatok a sejten belül, az organellumok membránjaiban is végbemennek. A különböző organellumok specifikus tartalma részben belső szintézissel, részben pedig a citoszolból történő aktív transzport révén jön létre. Ez utóbbira példa a fent említett Ca 2+ pumpa az izomsejtek szarkoplazmatikus retikulumában. Külön érdekesség, hogy a mitokondriumokban zajló ATP szintézis esetén a plazmamembrán ATPáz pumpáiban végbemenőre az ellenkező elv érvényes (1.6. ábra). Az ATP szintézis során az oxidatív anyagcsere meredek gradiens kialakulásához vezet H+ a belső membránokon. Ez a gradiens a mozgatórugója a molekulák aktív transzportjának fordított pumpálási ciklusának: a H + ionok a gradiens mentén mozognak a membránon, és az ennek eredményeként felszabaduló energia biztosítja az ATP szintézisét ADP-ből és foszfátból. A keletkező ATP pedig energiát ad a sejtnek, beleértve az aktív transzportot is.
Szállítás hólyagokban . A sejtben nagyszámú organellum és kapcsolódó hólyag található (1.1. ábra). Ezek az organellumok és különösen a vezikulák állandó mozgásban vannak, és tartalmukat más organellumokhoz vagy a plazmamembránhoz szállítják. A hólyagok a sejtmembránról az organellumokba is vándorolhatnak, mint az endocitózisban.
Folyamat fehérje szekrécióábrán látható. 1.12 A. A fehérje a sejtmag közelében szintetizálódik az endoplazmatikus retikulummal (az úgynevezett szemcsés, vagy durva, endoplazmatikus retikulummal) kapcsolatos riboszómákon; Az endoplazmatikus retikulumba jutva a fehérje transzportvezikulákba csomagolódik, amelyek elválik az organellumtól, és a Golgi-készülékbe vándorolnak. Itt egyesülnek a Golgi-készülék tartályaival, ahol a fehérje módosul (vagyis glikoproteinné alakul). A ciszternák végein a hólyagok ismét elválnak. A módosított fehérjét hordozó szekréciós vezikulák a plazmamembrán felé mozognak, és exocitózissal szabadítják fel tartalmukat.
Egy másik példa egy cellában lévő szállítási útvonalra az ábrán látható. 1,12, B; a koleszterin felvétele a sejt által. A vérben szállított koleszterin főként fehérjékhez, például részecskékhez kapcsolódik "alacsony sűrűségű lipoprotein"(LNP). Ezek a részecskék a membrán specifikus LDL receptor helyeihez kötődnek, ahol az endocitózis megtörténik, és az LDL "bevont" vezikulákban transzportálódik a sejtbe. Ezek a hólyagok endoszómákat képezve egyesülnek, és a folyamat során elveszítik „csengésüket”. Az endoszómák viszont fuzionálnak a túlnyomórészt hidrolitikus enzimeket tartalmazó primer lizoszómákkal, hogy másodlagos, nagyobb lizoszómákat képezzenek. Bennük a koleszterin felszabadul az LDL-részecskékből, és a citoszolba diffundál, ahol elérhetővé válik például a lipidmembránok szintéziséhez. Az LDL-t nem tartalmazó vezikulák is elválik az endoszómáktól, amelyek speciális módon a plazmamembránhoz vándorolnak, és azzal egyesülve visszaadják a membránanyagot és valószínűleg az LDL receptorokat. Attól a pillanattól kezdve, hogy az LDL részecske kötődik a membránhoz, 10-15 perc telik el, amíg a koleszterin felszabadul a másodlagos lizoszómából. Az LDL megkötésének és felszívódásának, azaz a sejt koleszterinnel való ellátásának károsodása döntő szerepet játszik egy súlyos és széles körben elterjedt betegség, az érelmeszesedés (artériák „megkeményedése”) kialakulásában.
Sok más szállítási útvonal is létezik, amelyek hasonlóak az ábrán láthatókhoz. 1.11 és 1.12,A, amelyek segítségével specifikus vezikulák mozognak a sejtben. Nem ismert, hogy pontosan hogyan mozognak, de valószínűleg a citoszkeleton elemei vesznek részt ebben a folyamatban. A vezikulák a mikrotubulusok mentén csúszhatnak, ilyenkor úgy tűnik, hogy a mozgáshoz szükséges energiát a hólyaghoz kapcsolódó fehérje, az ATPáz biztosítja (lásd alább). Teljesen érthetetlen marad, hogy hány különböző, minden irányban egymás után mozgó vezikula jut el a rendeltetési helyére. Ezeket nyilván úgy kell "megjelölni", hogy azt a közlekedési rendszer felismerje és célirányos mozgássá alakítsa.
Szállítás organellumok képződésével és elpusztításával . Eddig az endo- és exocitózist a vezikulák tartalmának szállításának folyamatának tekintettük. Van még egy aspektusa ezeknek a folyamatoknak, ami abban áll, hogy a plazmamembránnak a sejtfelszín egyik területén endocitózissal történő irányított eltávolítása, és éppen ellenkezőleg, exocitózissal egy másikhoz való hozzáadása, a sejtfelszín jelentős részeit megmozgatja. membrán (1.12.E ábra), lehetőséget adva a sejtnek például kinövés kialakítására vagy mozgásra.
Hasonló átrendeződések jellemzőek a citoszkeletonra is, különösen a mikrofilamentumokra és mikrotubulusokra (1.1. ábra). Mikrofilamentumok elsősorban abból állnak F-aktin fehérje amely a citoszolból származó monomer polimerizációja eredményeként rostos kötegekké képes összeállni. A kötegek polarizáltak, azaz gyakran csak az egyik végükről nőnek fel, új aktinmolekulákat halmoznak fel, míg a másik vége inert, vagy itt történik a szétszedés. Ennek a polarizált növekedésnek köszönhetően a mikrofilamentumok hatékonyan mozognak, és hálózatuk szerkezete megváltozhat. Az aktin átmenete depolimerizált állapotból (szol) szervezett állapotba (gél) nagyon gyorsan megtörténhet más fehérjék hatására vagy az ionkoncentráció változása alatt (lásd alább). Vannak olyan fehérjék is, amelyek hatására az aktin filamentumok rövid darabokra bomlanak. Számos sejt vékony kinövése - filopodia - tartalmaz egy központi aktinköteget (1.1. ábra), és a filopodiák különböző mozgásai valószínűleg az aktin átmenetei miatt következnek be: polimerizáció - depolimerizáció.
mikrotubulusokis gyakran esnek át hasonló mozgásokon. Ezeknek a mozgásoknak a mechanizmusa hasonló - a tubulin polimerizációja a citoszolból oly módon, hogy a mikrotubulus egyik vége megnő, míg a másik vagy nem változik, vagy ott szétesik. Így a mikrotubulus az anyag megfelelő hozzáadásával vagy eltávolításával át tud mozogni a citoszolon.
A citoszkeleton aktív mozgásai . A citoszkeletális struktúrák változásai mind az aktív mozgások, mind a fentebb leírt átrendeződések következtében felléphetnek. Sok esetben a mikrotubulusok és aktin filamentumok mozgását összehúzó fehérjék hajtják, amelyek megkötik a filamentumokat vagy tubulusokat, és el tudják mozgatni azokat egymáshoz képest. Mókusok miozin és dynein viszonylag magas koncentrációban jelen van az összes sejt citoszoljában; ezek azok az elemek, amelyek az energiát mozgássá alakítják a speciális sejtekben (izom) és organellumokban (csillók). Az izomsejtekben a miozin vastag filamentumokat képez, amelyek párhuzamosak az aktinszálakkal. A miozin molekula a „fejével” az aktin filamentumhoz kapcsolódik, és az ATP energiáját felhasználva kiszorítja a miozint az aktin molekula mentén. A miozin ezután leválik az aktinról. A sok ilyen kapcsolat-bontási ciklus halmaza makroszkopikushoz vezet izomrostok összehúzódása(4. fejezet). A dynein hasonló szerepet játszik a mikrotubulusok mozgásában a csillók működése során (1.1. ábra). A nem specializálódott sejtek citoplazmájában a miozin és a dynein nem szabályos rostokat, hanem a legtöbb esetben kis molekulacsoportokat alkot. Még ilyen kis aggregátumok formájában is képesek aktinszálakat vagy mikrotubulusokat mozgatni. Rizs. Az 1.13 ezt a folyamatot szemlélteti, amikor ellentétes polaritású miozinmolekulák is kapcsolódnak két különböző irányban polarizált aktinfilamentumhoz. A miozin fejcsoportjai a molekula farka felé hajlanak, ATP-t fogyasztanak, és a két aktin filamentum ellentétes irányba tolódik el, majd a miozin leválik róluk. Az ilyen jellegű mozgások, amelyek során az ATP energiája mechanikai munkává alakul át, megváltoztathatják a citoszkeleton és ennek következtében a sejtek alakját, valamint biztosíthatják a citoszkeletonhoz kapcsolódó organellumok szállítását.
Az intracelluláris transzport folyamatai legtisztábban egy idegsejt axonján mutathatók ki. axon transzport itt részletesen tárgyaljuk, hogy illusztráljuk azokat az eseményeket, amelyek valószínűleg a legtöbb sejtben hasonló módon fordulnak elő. A mindössze néhány mikron átmérőjű axon egy méter vagy több is lehet, és évekbe telne, amíg a fehérjék diffúzió útján eljutnak a magból az axon távolabbi végébe. Régóta ismert, hogy amikor az axon bármely szakasza szűkülésen megy keresztül, az axon proximális része kitágul. Úgy tűnik, hogy a centrifugális áramlás blokkolva van az axonban. Ilyen áramlás-gyors axontranszport képesábrán látható kísérlet szerint a radioaktív markerek mozgása bizonyítja. 1.14. Radioaktívan jelölt leucint injektáltunk a hátsó gyökér ganglionba, majd a 2. órától a 10. óráig radioaktivitást mértünk az ülőidegben a neurontestektől 166 mm távolságra. 10 órán keresztül a radioaktivitás csúcsa az injekció beadásának helyén nem változott jelentősen. De a radioaktivitás hulláma az axon mentén állandó sebességgel terjedt, körülbelül 34 mm/2 óra, vagyis 410 mm/nap. Kimutatták, hogy a homoioterm állatok minden neuronjában azonos ütemben megy végbe a gyors axontranszport, és nincs észrevehető különbség a vékony, nem myelinizált rostok és a legvastagabb axonok, valamint a motoros és szenzoros rostok között. A radioaktív marker típusa sem befolyásolja a gyors axontranszport sebességét; Különféle radioaktív anyagok szolgálhatnak markerként.

Rizs. 1.13.Egy bizonyos orientációjú, nem izomból álló miozin komplex különböző polaritású aktinszálakhoz kötődhet, és az ATP energiáját felhasználva eltolja azokat egymáshoz képest.
olyan molekulák, mint például a különböző aminosavak, amelyek beépülnek egy neuron testfehérjéibe. Ha az ideg perifériás részét elemezzük, hogy meghatározzuk az ide szállított radioaktivitás hordozóinak jellegét, akkor ilyen hordozók elsősorban a fehérjefrakcióban találhatók, de a mediátorok és a szabad aminosavak összetételében is. Tudva, hogy ezeknek az anyagoknak a tulajdonságai eltérőek, és különösen a molekuláik mérete eltérő, az állandó szállítási sebességet csak a mindegyikükre jellemző transzportmechanizmussal magyarázhatjuk.
Fentebb leírtuk gyors axontranszport van anterográd azaz a sejttesttől elfelé irányítva. Kimutatták, hogy egyes anyagok a perifériáról a sejttestbe jutnak a segítségével retrográd közlekedés. Például az acetil-kolinészteráz ebbe az irányba kétszer kisebb sebességgel szállítódik, mint a gyors axonális transzport sebessége. A neuroanatómiában gyakran használt marker – a torma-peroxidáz – szintén visszafelé mozog. A retrográd transzport valószínűleg fontos szerepet játszik a fehérjeszintézis szabályozásában a sejttestben. Néhány nappal az axon átvágása után kromatolízis figyelhető meg a sejttestben, ami a fehérjeszintézis megsértését jelzi. A kromatolízishez szükséges idő korrelál az axon transzekció helyétől a sejttestbe történő retrográd transzport időtartamával. Ez az eredmény magyarázatot is sugall erre a jogsértésre - a fehérjeszintézist szabályozó „jelanyag” perifériájáról történő átvitele megszakad. Nyilvánvaló, hogy a fő "járművek" használják a gyors axonális
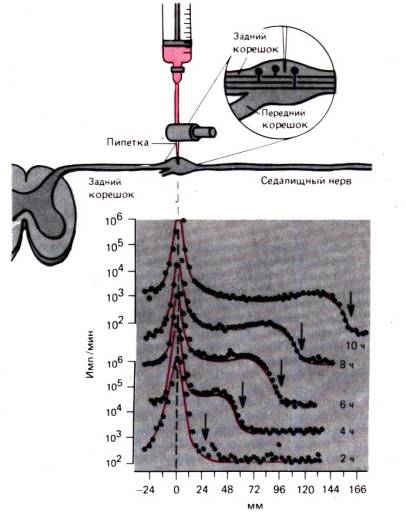
Rizs. 1.14.Kísérlet, amely egy macska ülőidegének szenzoros rostjaiban gyors axontranszportot mutat be. A tríciummal jelölt leucint a hátsó gyökér ganglionjába fecskendezik, és a ganglionban és a szenzoros rostokban lévő radioaktivitást 2, 4, 6, 8 és 10 órával az injekció beadása után mérik. (az ábra alján).Által abszcissza a gangliontól az ülőideg azon szakaszaiig való távolság, ahol a mérést végzik, elhalasztják. Az y tengelyen csak a felső és alsó görbék esetében logaritmikus skálán ábrázoljuk a radioaktivitást (imp./perc). A megnövekedett radioaktivitás "hulláma". (nyilak) 410 mm/nap sebességgel mozog (by )
a közlekedés hólyagok (vezikulák) és organellumok, mint például a szállítandó anyagokat tartalmazó mitokondriumok. A legnagyobb hólyagok vagy mitokondriumok mozgása mikroszkóppal figyelhető meg in vivo . Az ilyen részecskék rövid, gyors mozdulatokat tesznek egy irányba, megállnak, gyakran kissé hátra vagy oldalra mozdulnak, újra megállnak, majd egy szaggatást hajtanak végre a fő irányba. 410 mm/nap körülbelül 5 μm/s átlagos anterográd sebességnek felel meg; az egyes mozgások sebességének ezért sokkal nagyobbnak kell lennie, és ha figyelembe vesszük az organellumok, filamentumok és mikrotubulusok méretét, akkor ezek a mozgások valóban nagyon gyorsak. A gyors axontranszport jelentős ATP koncentrációt igényel. Az olyan mérgek, mint a mikrotubulusokat elpusztító kolhicin, szintén gátolják a gyors axontranszportot. Ebből az következik, hogy az általunk vizsgált transzportfolyamatban a vezikulák és organellumok mikrotubulusok és aktinszálak mentén mozognak; ezt a mozgást a dynein és a miozin molekulák kis aggregátumai biztosítják, amelyek az 1. ábrán látható módon hatnak. 1,13, az ATP energiáját felhasználva.
A gyors axontranszport is szerepet kaphat kóros folyamatok. Egyes neurotróp vírusok (például herpesz vagy gyermekbénulás vírusok) a periférián behatolnak az axonba, és retrográd transzport segítségével eljutnak a neurontestbe, ahol szaporodnak és kifejtik toxikus hatásukat. A bőrelváltozásokon keresztül a szervezetbe kerülő baktériumok által termelt fehérje, a tetanusztoxin az idegvégződéseken keresztül jut el az idegsejtekbe, ahol jellegzetes izomgörcsöket okoz. Magára az axontranszportra gyakorolt toxikus hatások esetei ismertek, például az akrilamid ipari oldószerrel való kitettsége. Ezenkívül úgy gondolják, hogy a beriberi beriberi és az alkoholos polyneuropathia patogenezise magában foglalja a gyors axontranszport megsértését.
A sejtben a gyors axontranszport mellett van egy meglehetősen intenzív lassú axontranszport. A tubulin körülbelül 1 mm/nap sebességgel mozog az axon mentén, míg az aktin gyorsabban, akár 5 mm/nap sebességgel mozog. Más fehérjék is vándorolnak a citoszkeleton ezen komponenseivel; például úgy tűnik, hogy az enzimek aktinhoz vagy tubulinhoz kapcsolódnak. A tubulin és az aktin mozgási sebessége nagyjából megegyezik a korábban leírt mechanizmus növekedési sebességével, amikor molekulákat építenek be egy mikrotubulus vagy mikrofilamentum aktív végébe. Ezért ez a mechanizmus állhat a lassú axontranszport hátterében. A lassú axontranszport sebessége hozzávetőlegesen megfelel az axonnövekedés sebességének is, ami nyilvánvalóan jelzi a citoszkeleton szerkezete által a második folyamatra szabott korlátokat.
A fejezet végén hangsúlyozni kell, hogy a sejtek semmiképpen sem statikus struktúrák, ahogyan például az elektronmikroszkópos fényképeken megjelennek. plazma membránés főleg az organellumok állandó gyors mozgásban és állandó átstrukturálódásban vannak; csak ezért képesek működni. Továbbá ezek nem egyszerű kamrák, amelyekben kémiai reakciók játszódnak le, hanem membránok és rostok magasan szervezett konglomerátumai, amelyben a reakciók optimálisan szervezett sorrendben mennek végbe.
1.4. A sejtfunkciók szabályozása
Az egyes sejt funkcionális egységként való fenntartását nagymértékben a sejtmag szabályozza; az ilyen szabályozó mechanizmusok tanulmányozása a sejtbiológia és a biokémia tárgya. Ugyanakkor a sejteknek a környezeti feltételeknek és a szervezet más sejtjeinek szükségleteinek megfelelően módosítaniuk kell működésüket, azaz funkcionális szabályozás tárgyaiként kell szolgálniuk. Az alábbiakban röviden áttekintjük, hogyan hatnak ezek a szabályozó hatások a plazmamembránra, és hogyan jutnak el az intracelluláris organellumokhoz.
Szabályozó hatások a sejtmembránra
Membránpotenciál . A sejtfunkciók szabályozása sok esetben a membránpotenciál változtatásával történik. Helyi potenciálváltozások akkor lehetségesek, ha: 1) a szomszédos sejtterületről vagy egy másik sejt által generált áram folyik át a membránon; 2) az ionok extracelluláris koncentrációja változik (gyakran [K + ] ki ); 3) megnyílnak a membrán ioncsatornái. A membránpotenciál változása befolyásolhatja a membránfehérjék konformációját, ami különösen a csatornák nyitását vagy zárását okozhatja. A fent leírtak szerint egyes membránszivattyúk működése a membránpotenciáltól függ. Az idegsejtek arra specializálódtak, hogy a membránpotenciál változásait feldolgozandó és továbbítandó információként érzékeljék (lásd a 2. fejezetet).
extracelluláris szabályozó anyagok . Az extracelluláris anyagokat érintő legfontosabb szabályozó mechanizmus a plazmamembránon vagy a sejten belüli specifikus receptorokkal való kölcsönhatásuk. Ezek az anyagok közé tartoznak a szinaptikus mediátorok, amelyek információt továbbítanak az idegsejtek között, helyi ágensek és olyan anyagok, amelyek a vérben keringenek, és elérik a test minden sejtjét, például hormonokat és antigéneket. szinaptikus neurotranszmitterek kis molekulák szabadulnak fel a szinapszisban lévő idegvégződésekből;
amikor elérik a szomszédos posztszinaptikus sejt plazmamembránját, elektromos jeleket vagy más szabályozó mechanizmusokat indítanak el. Ezt a kérdést részletesen a fejezet tartalmazza. 3.
Helyi vegyi anyagok gyakran speciális sejtek választják ki. Az extracelluláris térben szabadon diffundálnak, de hatásuk a sejtek egy kis csoportjára korlátozódik, mivel ezek az anyagok akár spontán, akár enzimek hatására gyorsan elpusztulnak. Az ilyen szerek felszabadulásának egyik példája a felszabadulás hisztamin hízósejtek sérülés vagy immunválasz esetén. A hisztamin ellazítja a vaszkuláris simaizomsejteket, növeli az erek endotéliumának permeabilitását, és stimulálja a viszketés érzését közvetítő érzőideg-végződéseket. Más helyi vegyi anyagokat sok más sejt választ ki. Tipikus helyi ügynökök prosztaglandinok, körülbelül 20 zsírsavszármazékból álló csoportot alkotnak. A széles körben elterjedt sejtekből folyamatosan szabadulnak fel, de csak lokálisan fejtik ki hatásukat, mivel a membránfoszfolipázok gyorsan elpusztítják őket. A különféle prosztaglandinok széles hatásspektrummal rendelkeznek: kiválthatják a simaizomsejtek összehúzódását, a vérlemezkék (thrombocyták) aggregációját okozhatják, vagy gátolhatják a sárgatest fejlődését a petefészekben.
Más helyi ügynökök szolgálnak növekedési tényezők. A szimpatikus idegsejtek legismertebb idegnövekedési faktora (NGF), amely szükséges ezen idegsejtek növekedéséhez és túléléséhez a fejlődés során in vivo vagy sejtkultúrában. Nyilvánvaló, hogy a neuronok ezen osztályának célsejtjei NGF-et választanak ki, és így biztosítják a megfelelő beidegzést. A szervek kialakításakor a sejteknek gyakran „meg kell találniuk az utat” a célsejtekhez, amelyek jelentős távolságra helyezkedhetnek el. Ennek megfelelően sok speciális növekedési faktornak kell lennie, mint például az NGF.
Hormonok és antigének a vér minden sejthez eljuttatja. Az antigének immunválaszt váltanak ki a specifikus antitesteket hordozó sejtekből. Az antigének azonban általában olyan idegen anyagok, amelyek nem képződnek a reagáló szervezetben (további részletekért lásd a 18. fejezetet). Egyes hormonok, például az inzulin vagy a tiroxin a sejttípusok széles skáláját érintik, míg mások, például a nemi hormonok csak bizonyos sejttípusokra. A hormonok vagy peptidek, amelyek hatását a sejtmembránon lévő receptorokhoz való kötődésük váltja ki, vagy szteroidok és tiroxin, amelyek a lipidmembránon keresztül diffundálnak és az intracelluláris receptorokhoz kötődnek. A szteroid hormonok a nukleáris kromatinhoz kötődnek, ami bizonyos gének átírását eredményezi. Az ennek eredményeként keletkező fehérjék a sejtek működésében változást okoznak, ami a hormonok specifikus hatása. A hormonok felszabadulásával és hatásával kapcsolatos kérdéseket a fejezet részletesen tárgyalja. 17.
Intracelluláris kommunikáció második hírvivők bevonásával
A fent leírt szabályozó funkciók közé tartozik a sejtmembránra gyakorolt hatás. A sejtmembrán által kapott információnak gyakran reakciót kell okoznia az organellumokban, és különféle anyagok, másodlagos hírvivők (szemben az elsővel, amelyek külső forrásból jutnak a sejtbe) eljuttatják hozzájuk. A második közvetítők tanulmányozása gyorsan fejlődik, és nincs garancia arra, hogy a probléma jelenlegi megértési szintje kellően teljes lesz. Itt három jól tanulmányozott mediátort érintünk: Ca 2+ , cAMP és inozitol-trifoszfát.
Kalcium.A legegyszerűbb intracelluláris mediátor a Ca 2+ -ion. Szabad koncentrációja nyugvó sejtben nagyon alacsony, 10 -8 -10 -7 mol/l. Meghatározott membráncsatornákon keresztül juthat be a sejtbe, ha azok nyitva vannak, például amikor a membránpotenciál megváltozik (lásd 2. fejezet). Az ebből eredő Ca 2+ növekedése fontos reakciókat indít el a sejtben, mint például az izomösszehúzódás alapját jelentő myofibrillumok összehúzódása (lásd 4. fejezet), vagy neurotranszmitter tartalmú vezikulák felszabadulása az idegvégződésekből (lásd 3. fejezet). . Mindkét reakcióhoz körülbelül 10-5 mol/l Ca 2+ -koncentráció szükséges. A szabályozó hatású Ca 2+ az intracelluláris depóból is felszabadulhat, például az endoplazmatikus retikulumból. A Ca 2+ raktárból történő felszabadítása más közvetítők részvételét igényli (lásd pl. 1.16. ábra).
Ciklikus adenozin-monofoszfát, cAMP. A közelmúltban bebizonyosodott, hogy a ciklikus adenozin-monofoszfát (cAMP), a szervezet fő energiaforrásának, az ATP-nek a származéka, fontos másodlagos hírvivő. ábrán látható összetett reakciólánc. 1,15, a receptortól indul Rs a plazmamembrán külső felületén, amely specifikus kötőhelyként szolgálhat a különböző mediátorok és hormonok számára. Egy specifikus "stimuláló" molekulához való kötődés után Rs megváltoztatja a felépítését; ezek a változások befolyásolják a fehérjét Gs a membrán belső felületén oly módon, hogy lehetővé válik az utóbbi aktiválása intracelluláris guanozin-trifoszfáttal (GTP). aktivált fehérje Gs , viszont serkenti a membrán belső felületén található enzimet, az adenilát-ciklázt (AC), amely katalizálja a cAMP képződését az ATP-ből. Vízben oldódó cAMP és a közvetítő, amely továbbítja a hatást

Rizs. 1.15.A cAMP (ciklikus adenozin-monofoszfát) intracelluláris mediátort tartalmazó reakciólánc. A serkentő vagy gátló külső jelek aktiválják az R membránreceptorokat s vagy Ri . Ezek a receptorok szabályozzák a kötődési folyamatot G - fehérjék intracelluláris GTP-vel (guanozin-trifoszfát), ezáltal stimulálják vagy gátolják az intracelluláris adenilát-ciklázt (AC). Az AC amplifikáló enzim az adenozin-trifoszfátot (ATP) cAMP-vé alakítja, amely aztán foszfodiészter segítségével AMP-vé hasad. A szabad cAMP bediffundál a sejtbe és aktiválja az adenilát kinázt (A-kináz), felszabadítva annak katalitikus C alegységét, amely katalizálja az intracelluláris fehérjék foszforilációját, azaz. képezi az extracelluláris inger végső hatását. A séma olyan farmakológiai gyógyszereket és toxinokat is bemutat, amelyek bizonyos reakciókat váltanak ki (+) vagy gátolnak (-) (de változásokkal)
egy extracelluláris receptor stimulálása Rs a sejt belső struktúráihoz.
Párhuzamosan a stimuláló reakciólánccal, amely magában foglalja Rs gátló mediátorok és hormonok lehetséges kötődése a megfelelő receptorhoz R i amely ismét GTP-aktivált fehérjén keresztül G , gátolja az AC és így a cAMP termelődését. A sejtbe diffundálva a cAMP reakcióba lép az adenilát-kinázzal (A-kináz); Ez felszabadítja a C alegységet, amely katalizálja a fehérje foszforilációját. Ez a foszforiláció a fehérjéket aktív formájukká alakítja, és most már kifejthetik sajátos szabályozó hatásukat (például glikogén lebomlását). Ez a komplex szabályozórendszer rendkívül hatékony, hiszen a végeredmény számos fehérje foszforilációja, azaz a szabályozó jel nagy amplifikációs faktorral halad át az áramkörön. Külső mediátorok, amelyek a receptorokhoz kötődnek R s és R i mindegyikre jellemző rendkívül változatos. Adrenalin, kavar R s vagy R i részt vesz a lipid és glikogén anyagcsere szabályozásában, valamint a szívizom fokozott összehúzódásában és egyéb reakciókban (lásd 19. fejezet). pajzsmirigy-stimuláló hormon, aktiválása Rs , serkenti a tiroxin hormon szekrécióját a pajzsmirigy által, a prosztaglandin I pedig gátolja a vérlemezkék aggregációját. A gátló hatások, beleértve az adrenalint is, közvetítik R i a lipolízis lassításában fejeződik ki. És így, a cAMP rendszer egy többfunkciós intracelluláris szabályozó rendszer, amely extracelluláris stimuláló és gátló jelátviteli ágensekkel pontosan szabályozható.
Inozitol-foszfát "IF h ". A második mediátor, az inozitol-foszfát intracelluláris rendszerét csak nemrég fedezték fel (1.16. ábra). Ebben az esetben nincs gátló út, de van hasonlóság a cAMP rendszerrel, amelyben az R receptor stimulálásának hatása átkerül a membrán belső felületén lévő GTP-aktivált G-fehérjére. A következő lépésben a szokásos membránlipid foszfatidil-inozitol (PI), amely korábban két további foszfátcsoportot kapott, PI-difoszfáttá (FIF 2) alakul, amelyet az aktivált foszfodiészteráz (PDE) hasít. inozitol-trifoszfát(IFz) és lipid diacil-glicerin(DAG). Az inozitol-trifoszfát egy vízben oldódó másodlagos hírvivő, amely a citoszolba diffundál. Elsősorban Ca 2+ felszabadításával fejti ki hatását az endoplazmatikus retikulumból. A Ca 2+ viszont a fent leírtak szerint közvetítőként működik; például aktiválja a Ca2+-függő foszfokinázt, amely foszforilálja az enzimeket. A DAG lipid alegysége (1.16. ábra) szintén a plazmamembrán lipid fázisában diffúzióval továbbítja a jelet a belső felületén található C-kináz felé, amely a foszfatidil-szerin, mint kofaktor részvételével aktiválódik. Ezután a C-kináz kiváltja a fehérjék foszforilációját, aktív formává alakítva azokat.
A második intermedier 3 interferon intracelluláris rendszerét számos külső mediátor és hormon szabályozhatja, beleértve az acetilkolint, a szerotonint, a vazopresszint és a pajzsmirigy-stimuláló hormont; a cAMP rendszerhez hasonlóan sokféle intracelluláris hatás jellemzi. Lehetséges, hogy ezt a rendszert a szem vizuális receptorában lévő fény is aktiválja, és központi szerepet játszik a fototranszdukcióban (lásd a 11. fejezetet). A szervezet egyedfejlődése során először aktiválják az IGF-rendszer receptorát a spermiumok, melynek eredményeként az IGF részt vesz a petesejt megtermékenyítését kísérő szabályozási reakciókban.
A cAMP és IFz-DAG rendszerek rendkívül hatékonyak biológiai fokozók.Ők

Rizs. 1.16.Az intracelluláris intermedier interferont (inozitol-trifoszfátot) tartalmazó reakciólánc. A cAMP rendszerhez hasonlóan az extracelluláris jelet egy fehérje közvetítiG, amely ebben az esetben a foszfodiészterázt (PDE) aktiválja. Ez az enzim lebontja a foszfatidilinozin-difoszfátot (FIF) 2 ) a plazmamembránban az IF előtt h és diacil-glicerin (DAG); HA h diffundál a citoplazmába. Itt okozza a Ca felszabadulását 2+ az endoplazmatikus retikulumból; Ca-koncentráció növekedése 2+ a citoplazmában ([Ca 2+ ]i ) aktivál egy protein-kinázt, amely foszforilál, és ezáltal aktiválja az enzimeket. Egy másik termék, a DAG, a membránban marad, és aktiválja a protein-kináz C-t (kofaktor-foszfatidil-szerin, PS). A protein-kináz C olyan enzimeket is foszforilál, amelyek a külső receptor stimulálásával kapcsolatos specifikus hatásokat közvetítik. R . Az IF-t érintő reakciólánc ágai h és a DAG egymástól függetlenül aktiválható ionomicinnel és forbol-észterrel (módosítva)
A mediátor és a külső membránreceptor közötti reakciót számos intracelluláris fehérje foszforilációjává alakítják át, amelyek azután különböző sejtfunkciókat befolyásolhatnak. A probléma egyik lényeges aspektusa, hogy mai ismereteink szerint csak ez a két, egymással szorosan összefüggő ilyen típusú szabályozórendszer létezik, amelyeket számos külső mediátor használ különböző intracelluláris folyamatok szabályozására. Ugyanakkor ezek a szabályozó rendszerek, beleértve a Ca 2+ -ot is, szoros kölcsönhatásban állnak egymással, ami lehetővé teszi számukra a sejtfunkciók finom szabályozását.
Oktatóanyagok és útmutatók
1. Alberts BAN BEN., Bray D., Lewis J., Raff M., Roberts NAK NEK., Watson J.D.Molecular Biology of the Cell, New York és London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(szerk.). Biológia. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille BAN BEN. Gerjeszthető membránok ioncsatornái. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(szerk.). Biophysik. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann NAK NEK., Mahler H.Biokémia. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E.R., Schwartz-J.H.,(szerk.). Az idegtudomány alapelvei, New York, Amszterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., SchmidtW.Anatómiai des Menschen. Berlin, Heidelberg, New York, Tokió, Springer, 1983.
Eredeti cikkek és vélemények
8. Berridge M.J. A sejten belüli kommunikáció molekuláris alapja, Sci. Amer 253, 124, 134 (1985).
9. Berridge M. J., Irvine R. F. Inozitol-trifoszfát, egy új második hírvivő a sejtes jelátvitelben. Nature, 312, 315, 321 (1984).
10. Bretscher M.S. A sejtmembrán molekulái, Sci. Amer., 253, 124–134 (1985).
11. Daut J. Az élő sejt mint energiaátalakító gép. A szívizom metabolizmusának minimális modellje, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz BAN BEN. A nátriumionok hatása a tintahal óriás axonjának elektromos aktivitására. J Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Kationok aktív transzportja óriási axonokban innen Szépiaés loligo, J Physiol. (Lond.), 128, 28–42 (1955).
14. Hosszabb P. Ioncsatornák konformációs részállapotokkal, Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M. Exoplazmatikus transzport normál és patológiás rendszerekben. In: Physiology and Pathology of Axons, S.G. Waxman, Szerk. New York, Raven Press, 1978.
Meghívjuk Önt, hogy ismerkedjen meg az anyagokkal és.
: cellulóz membrán, membrán, citoplazma sejtszervekkel, sejtmag, vakuolák sejtnedvvel.A plasztidok jelenléte a növényi sejt fő jellemzője.
A sejtfal funkciói- meghatározza a sejt alakját, véd a környezeti tényezők ellen.
plazma membrán- vékony film, kölcsönhatásban lévő lipid- és fehérjemolekulákból áll, elhatárolja a belső tartalmat a külső környezettől, ozmózissal és aktív transzferrel biztosítja a víz, ásványi és szerves anyagok sejtbe szállítását, valamint eltávolítja a salakanyagokat.
Citoplazma- a sejt belső félfolyékony környezete, amelyben a sejtmag és az organellumok találhatók, kapcsolatot biztosít közöttük, részt vesz az élet főbb folyamataiban.
Endoplazmatikus retikulum- elágazó csatornák hálózata a citoplazmában. Részt vesz a fehérjék, lipidek és szénhidrátok szintézisében, az anyagok szállításában. Riboszómák - az EPS-en vagy a citoplazmában található testek, RNS-ből és fehérjéből állnak, részt vesznek a fehérjeszintézisben. Az EPS és a riboszómák egyetlen berendezés a fehérjék szintézisére és szállítására.
Mitokondriumok- a citoplazmától két membránnal elválasztott organellumok. Szerves anyagok oxidálódnak bennük, és enzimek részvételével ATP-molekulák szintetizálódnak. A belső membrán felületének növekedése, amelyen az enzimek találhatók a cristae miatt. Az ATP egy energiában gazdag szerves anyag.
plasztidok(kloroplasztiszok, leukoplasztok, kromoplasztok), ezek sejttartalma a növényi szervezet fő jellemzője. A kloroplasztok olyan plasztiszok, amelyek zöld pigment klorofillt tartalmaznak, amely elnyeli a fényenergiát, és azt szerves anyagok szintetizálására használja fel szén-dioxidból és vízből. A kloroplasztok elhatárolása a citoplazmából két membránnal, számos kinövés - a belső membránon lévő grána, amelyben klorofillmolekulák és enzimek találhatók.
Golgi komplexus- a citoplazmától membránnal határolt üregrendszer. A fehérjék, zsírok és szénhidrátok felhalmozódása bennük. Zsírok és szénhidrátok szintézisének megvalósítása a membránokon.
Lizoszómák- a citoplazmától egyetlen membránnal elválasztott testek. A bennük lévő enzimek felgyorsítják a komplex molekulák egyszerű felosztásának reakcióját: a fehérjéket aminosavakra, az összetett szénhidrátokat az egyszerűekre, a lipideket a glicerinre és a zsírsavakra, valamint elpusztítják a sejt elhalt részeit, az egész sejteket.
Vacuolák- sejtnedvvel töltött üregek a citoplazmában, tartalék tápanyagok, káros anyagok felhalmozódásának helye; szabályozzák a sejt víztartalmát.
Mag- a sejt fő része, kívülről kétmembránnal borított, pórusokkal áttört magburok. Az anyagok bejutnak a magba, és a pórusokon keresztül távoznak onnan. A kromoszómák örökletes információk hordozói egy szervezet jellemzőiről, a sejtmag fő szerkezeteiről, amelyek mindegyike egy DNS-molekulából és fehérjékkel kombinálva áll. A sejtmag a DNS, i-RNS, r-RNS szintézisének helye.

Külső membrán jelenléte, citoplazma organellákkal, mag kromoszómákkal.
Külső vagy plazmamembrán- elhatárolja a sejt tartalmát a környezettől (más sejtek, sejtközi anyag), lipid- és fehérjemolekulákból áll, biztosítja a sejtek közötti kommunikációt, az anyagok sejtbe történő szállítását (pinocitózis, fagocitózis) és a sejtből kifelé.
Citoplazma- a sejt belső félfolyékony környezete, amely kommunikációt biztosít a sejtmag és a benne található organellumok között. A létfontosságú tevékenység fő folyamatai a citoplazmában zajlanak.
Sejtorganellumok:
1) endoplazmatikus retikulum (ER)- elágazó tubulusok rendszere, amely részt vesz a fehérjék, lipidek és szénhidrátok szintézisében, az anyagok szállításában a sejtben;
2) riboszómák- az rRNS-t tartalmazó testek az ER-en és a citoplazmában helyezkednek el, és részt vesznek a fehérjeszintézisben. Az EPS és a riboszómák egyetlen berendezés a fehérjeszintézishez és -szállításhoz;
3) mitokondriumok- a sejt "erőművei", amelyeket két membrán határol el a citoplazmától. A belső cristae-kat (redőket) képez, amelyek megnövelik a felületét. A cristae-n lévő enzimek felgyorsítják a szerves anyagok oxidációs reakcióit és az energiadús ATP molekulák szintézisét;
4) golgi komplexum- a citoplazmából membránnal határolt üregek csoportja, amelyet fehérjékkel, zsírokkal és szénhidrátokkal töltenek meg, amelyeket vagy az életfolyamatok során használnak fel, vagy eltávolítanak a sejtből. A komplex membránjai zsírok és szénhidrátok szintézisét végzik;
5) lizoszómák- az enzimekkel feltöltött testek felgyorsítják a fehérjék aminosavakra, a lipidek glicerinre és zsírsavakra, a poliszacharidok monoszacharidokra történő hasadási reakcióit. A lizoszómákban a sejt elhalt részei, az egész sejtek és a sejtek elpusztulnak.
Sejtzárványok- Tartalék tápanyagok felhalmozódása: fehérjék, zsírok és szénhidrátok.
Mag- a sejt legfontosabb része. Kettős membrános membrán borítja, pórusokkal, amelyeken keresztül egyes anyagok behatolnak a sejtmagba, míg mások a citoplazmába. A kromoszómák a mag fő szerkezetei, az élőlény jellemzőiről szóló örökletes információ hordozói. Az anyasejt osztódása során a leánysejtekre, a csírasejtekkel pedig a leányszervezetekre terjed. A sejtmag a DNS, mRNS, rRNS szintézis helye.
Gyakorlat:
Magyarázza meg, miért nevezik az organellumokat a sejt speciális struktúráinak?
Válasz: Az organellumokat speciális sejtstruktúráknak nevezik, mivel szigorúan meghatározott funkciókat látnak el, az örökletes információkat a sejtmagban tárolják, az ATP-t a mitokondriumokban szintetizálják, a fotoszintézis a kloroplasztiszokban megy végbe stb.
Ha kérdése van a citológiával kapcsolatban, kérhet segítséget
Az evolúció harmadik szakasza a sejt megjelenése.
A fehérjék és nukleinsavak (DNS és RNS) molekulái biológiai sejtet, az élet legkisebb egységét alkotják. A biológiai sejtek minden élő szervezet "építőkövei", és tartalmazzák a fejlődés összes anyagi kódját.
A tudósok sokáig rendkívül egyszerűnek tartották a sejt szerkezetét. A szovjet enciklopédikus szótár a sejt fogalmát a következőképpen értelmezi: "A sejt egy elemi élő rendszer, minden állat és növény szerkezetének és életének alapja." Meg kell jegyezni, hogy az „elemi” kifejezés semmiképpen sem azt jelenti, hogy „a legegyszerűbb”. Ellenkezőleg, a sejt Isten egyedi fraktálalkotása, amely lenyűgöző összetettségében és egyúttal rendkívüli koherenciájában is. minden elemének munkája.
Amikor egy elektronmikroszkóp segítségével sikerült benéznünk a belsejébe, kiderült, hogy a legegyszerűbb sejt eszköze éppoly összetett és felfoghatatlan, mint maga az Univerzum. Ma már megállapították, hogy "A sejt az Univerzum különleges anyaga, a Kozmosz különleges anyaga." Egyetlen cella olyan információt tartalmaz, amely a Nagy Szovjet Enciklopédia néhány tízezer kötetébe helyezhető be. Azok. a sejt többek között az információ hatalmas "biotározója".
A modern molekuláris evolúciós elmélet szerzője, Manfred Eigen ezt írja: „Ahhoz, hogy egy fehérjemolekula véletlenül kialakuljon, a természetnek körülbelül 10130 próbát kellene elvégeznie, és erre annyi molekulát kellene költenie, hogy 1027-re elegendő lenne. Univerzumok. Ha a fehérjét intelligensen építették fel, vagyis hogy minden egyes lépés érvényességét valamilyen szelekciós mechanizmussal ellenőrizni lehessen, akkor ehhez mindössze körülbelül 2000 próbálkozás kellett volna. Paradox következtetésre jutunk: a "primitív élő sejt" felépítésére szolgáló program. valahol az elemi részecskék szintjén van kódolva".
És hogyan is lehetne másként. Minden sejt, amely DNS-sel rendelkezik, tudattal rendelkezik, tudatában van önmagának és más sejteknek, és kapcsolatban van az univerzummal, lévén annak része. És bár az emberi test sejtjeinek száma és változatossága elképesztő (körülbelül 70 billió), mindegyik önhasonló, ahogy a sejtekben végbemenő összes folyamat önhasonló. Roland Glaser német tudós szavai szerint a biológiai sejtek tervezése "nagyon jól átgondolt". Ki van jól átgondolva?
A válasz egyszerű: a fehérjék, nukleinsavak, élő sejtek és minden biológiai rendszer az intellektuális Teremtő kreatív tevékenységének termékei.
Ami érdekes: atomi szinten nincs különbség a szerves és szervetlen világ kémiai összetétele között. Más szóval, az atom szintjén egy sejt ugyanazokból az elemekből jön létre, mint az élettelen természet. A különbségek molekuláris szinten találhatók. Az élő testekben a szervetlen anyagok és a víz mellett fehérjék, szénhidrátok, zsírok, nukleinsavak, ATP-szintáz enzim és egyéb kis molekulatömegű szerves vegyületek is megtalálhatók.
A mai napig a sejtet szó szerint atomokra bontották tanulmányozás céljából. Azonban nem lehet létrehozni legalább egy élő sejtet, mert egy sejt létrehozása az élő Univerzum egy részecskéjének létrehozását jelenti. akadémikus V.P. Kaznacsejev úgy véli, hogy "a sejt egy kozmikus-bolygós organizmus... Az emberi sejtek bizonyos éter-torziós bioütköztető rendszerek. Ezekben a bioütközőkben számunkra ismeretlen folyamatok mennek végbe, kozmikus áramlási formák materializálódnak, kozmikus átalakulásuk megy végbe. , és ennek köszönhetően a részecskék materializálódnak" .
Víz.
A sejttömeg közel 80%-a víz. A biológiai tudományok doktora, S. Zenin szerint a víz klaszterszerkezeténél fogva a biokémiai folyamatok irányításának információs mátrixa. Ezenkívül a víz az elsődleges „célpont”, amellyel a hangfrekvenciás rezgések kölcsönhatásba lépnek. A sejtvíz rendezettsége olyan magas (közel a kristály rendezettségéhez), hogy folyadékkristálynak nevezik.
Mókusok.
A fehérjék fontos szerepet játszanak a biológiai életben. A sejt több ezer olyan fehérjét tartalmaz, amelyek csak erre a sejttípusra jellemzőek (az őssejtek kivételével). A saját fehérjék szintetizálásának képessége sejtről sejtre öröklődik, és az egész életen át fennmarad. A sejt élete során a fehérjék fokozatosan megváltoztatják szerkezetüket, működésük károsodik. Ezeket az elhasznált fehérjéket eltávolítják a sejtből, és újakkal helyettesítik, aminek köszönhetően a sejt élettevékenysége megmarad.
Mindenekelőtt a fehérjék építő funkcióját jegyezzük meg, mivel ezek a sejtek membránjait és sejtszervecskéit, az érfalakat, inakat, porcokat stb.
A fehérjék jelátviteli funkciója rendkívül érdekes. Kiderült, hogy a fehérjék képesek jelzőanyagként szolgálni, jeleket továbbítva szövetek, sejtek vagy organizmusok között. A jelátviteli funkciót hormonfehérjék látják el. A sejtek az intercelluláris anyagon keresztül továbbított jelátviteli fehérjék segítségével távolról tudnak kommunikálni egymással.
A fehérjéknek motoros funkciójuk is van. Mindenféle mozgást, amelyre a sejtek képesek, mint például az izomösszehúzódást, speciális kontraktilis fehérjék hajtják végre. A fehérjék szállítási funkciót is ellátnak. Különféle anyagokat képesek megkötni és a sejt egyik helyéről a másikra átvinni. Például a hemoglobin vérfehérje oxigént köt, és a test minden szövetébe és szervébe szállítja. Ezenkívül a fehérjéknek védő funkciójuk is van. Amikor idegen fehérjéket vagy sejteket juttatnak be a szervezetbe, abban speciális fehérjék képződnek, amelyek megkötik és semlegesítik az idegen sejteket, anyagokat. És végül a fehérjék energiafunkciója az, hogy 1 g fehérje teljes lebontásával 17,6 kJ energia szabadul fel.
Sejtszerkezet.
A sejt három elválaszthatatlanul összefüggő részből áll: membránból, citoplazmából és sejtmagból, és a sejtmag felépítése és működése a sejt életének különböző időszakaiban eltérő. Egy sejt élete ugyanis két időszakot foglal magában: osztódást, amelynek eredményeként két leánysejt képződik, és az osztódások közötti időszakot, amelyet interfázisnak neveznek.
A sejtmembrán közvetlenül kölcsönhatásba lép a külső környezettel és kölcsönhatásba lép a szomszédos sejtekkel. Egy külső rétegből és egy alatta elhelyezkedő plazmamembránból áll. Az állati sejtek felszíni rétegét glikokalisztnak nevezik. Összeköti a sejteket a külső környezettel és az azt körülvevő összes anyaggal. Vastagsága kisebb, mint 1 mikron.
Sejtszerkezet
A sejtmembrán nagyon fontos része a sejtnek. Összetartja az összes sejtkomponenst, és elhatárolja a külső és belső környezetet.
A sejtek és a külső környezet között állandó anyagcsere zajlik. A külső környezetből víz, különféle sók egyedi ionok formájában, szervetlen és szerves molekulák jutnak be a sejtbe. Az anyagcseretermékek, valamint a sejtben szintetizált anyagok: fehérjék, szénhidrátok, hormonok, amelyek különböző mirigyek sejtjeiben keletkeznek, a sejtből a membránon keresztül ürülnek ki a külső környezetbe. Az anyagok szállítása a plazmamembrán egyik fő funkciója.
Citoplazma- belső félfolyékony közeg, amelyben a fő anyagcsere folyamatok zajlanak. A legújabb tanulmányok kimutatták, hogy a citoplazma nem egyfajta megoldás, amelynek összetevői véletlenszerű ütközések során kölcsönhatásba lépnek egymással. A zseléhez hasonlítható, amely a külső hatásokra reagálva "remegni" kezd. A citoplazma így érzékeli és továbbítja az információt.
A sejtmag és a különféle organellumok a citoplazmában helyezkednek el, amelyeket ez egy egésszé egyesít, amely biztosítja kölcsönhatásukat és a sejt egyetlen integrált rendszerként való tevékenységét. A sejtmag a citoplazma központi részén található. A citoplazma teljes belső zónáját az endoplazmatikus retikulum tölti ki, amely sejtorganoid: tubulusokból, hólyagokból és "ciszternákból" álló rendszer, amelyet membránok határolnak. Az endoplazmatikus retikulum részt vesz az anyagcsere folyamatokban, biztosítja az anyagok szállítását a környezetből a citoplazmába és az egyes intracelluláris struktúrák között, de fő funkciója a fehérjeszintézisben való részvétel, amely a riboszómákban megy végbe. - 15-20 nm átmérőjű, kerek alakú mikroszkopikus kis testek. A szintetizált fehérjék először az endoplazmatikus retikulum csatornáiban és üregeiben halmozódnak fel, majd eljutnak az organellumokhoz és a sejthelyekhez, ahol elfogyasztják őket.
A citoplazma a fehérjéken kívül mitokondriumokat is tartalmaz, 0,2-7 mikron méretű kis testeket, amelyeket a sejtek "erőműveinek" neveznek. A redox reakciók a mitokondriumokban mennek végbe, amelyek energiával látják el a sejteket. A mitokondriumok száma egy sejtben néhánytól több ezerig terjed.
Mag- a sejt létfontosságú része, szabályozza a fehérjék szintézisét és ezen keresztül a sejtben zajló összes élettani folyamatot. A nem osztódó sejt magjában megkülönböztetik a magmembránt, a magnedvet, a sejtmagot és a kromoszómákat. A magburkon keresztül folyamatos anyagcsere megy végbe a sejtmag és a citoplazma között. A nukleáris burok alatt - maglé (félig folyékony anyag), amely tartalmazza a magot és a kromoszómákat. A nucleolus egy sűrű, lekerekített test, amelynek méretei nagyon eltérőek lehetnek, 1-10 mikron és több. Főleg ribonukleoproteinekből áll; részt vesz a riboszómák képződésében. Általában 1-3 sejtmag található, néha akár több száz is. A sejtmag RNS-ből és fehérjéből áll.
A sejt megjelenésével élet keletkezett a Földön!
Folytatjuk...


