Zagotavlja odnos celice z okoljem. Povezava organizma z okoljem
Povezava organizma z okoljem je s fizikalno-kemijskega vidika odprt sistem, torej sistem, kjer potekajo biokemični procesi. Začetne snovi prihajajo iz okolja, snovi, ki prav tako kontinuirano nastajajo, pa se izločajo. Ravnovesje med hitrostjo in koncentracijo produktov večsmernih reakcij v telesu je pogojno, namišljeno, saj se vnos in odstranjevanje snovi ne ustavi. Nenehna povezava z okoljem in nam omogoča, da živi organizem obravnavamo kot odprt sistem.
Sonce je vir energije za vse žive celice. Rastlinske celice zajamejo energijo sončne svetlobe s pomočjo klorofila in jo uporabljajo za asimilacijske reakcije med fotosintezo. Celice živali, gliv, bakterij uporabljajo sončno energijo posredno, ko delijo organske snovi, ki jih sintetizira zemeljska rastlina.
Del hranilnih snovi v celici se razgradi v procesu celičnega dihanja in tako oskrbi energijo, potrebno za različne vrste celične aktivnosti. Ta proces poteka v organelah, imenovanih mitohondriji. Mitohondrijo sestavljata dve membrani: zunanja, ki ločuje organele od citoplazme, in notranja, ki tvori številne gube. Glavni produkt dihanja je ATP. Zapusti mitohondrije in se uporablja kot vir energije za številne kemične reakcije v citoplazmi in celični membrani. Če je za izvajanje celičnega dihanja potreben kisik, potem dihanje imenujemo aerobno dihanje, če pa reakcije potekajo v odsotnosti kisika, potem govorimo o anaerobnem dihanju.
Za kakršno koli delo, opravljeno v celici, se energija uporablja v eni sami obliki – v obliki energije iz fosfatnih vezi ATP. ATP je zelo mobilna spojina. Tvorba ATP se pojavi na notranji membrani mitohondrijev. ATP se sintetizira v vseh celicah med dihanjem zaradi energije oksidacije ogljikovih hidratov, maščob in drugih organskih snovi. V celicah zelenih rastlin se glavna količina ATP sintetizira v kloroplastih zaradi sončne energije. V njih se med fotosintezo proizvede večkrat več ATP kot v mitohondrijih. ATP se razgradi s pretrganjem vezi fosfor-kisik in sproščanjem energije. To se zgodi pod delovanjem encima ATPaze v procesu hidrolize ATP - dodajanje vode z izločanjem molekule fosforne kisline. Posledično se ATP pretvori v ADP, in če se dve molekuli fosforne kisline odcepi, potem v AMP. Reakcijo cepitve vsake gram-molekule kisline spremlja sproščanje 40 kJ. To je zelo velik energetski donos, zato se vezi fosfor-kisik ATP običajno imenujejo makroergične (visokoenergijske).
Uporaba ATP v reakcijah plastične izmenjave se izvaja z njihovo konjugacijo s hidrolizo ATP. Molekule različnih snovi se z energijo napolnijo tako, da na molekulo ATP, torej s fosforilacijo, vežejo fosforno skupino, ki se sprosti med hidrolizo.
Značilnost fosfatnih derivatov je, da ne morejo zapustiti celice, čeprav njihove "izpraznjene" oblike prosto prehajajo skozi membrano. Zaradi tega fosforilirane molekule ostanejo v celici, dokler se ne uporabijo v ustreznih reakcijah.
Obratni proces pretvorbe ADP v ATP se zgodi tako, da na ADP pritrdimo molekulo fosforne kisline, pri čemer se sprosti voda in absorbira velika količina energije.
Tako je ATP univerzalni in neposredni vir energije za celično aktivnost. To ustvarja enoten celični sklad energije in omogoča njeno prerazporeditev in transport iz enega dela celice v drugega.
Prenos fosfatne skupine ima pomembno vlogo pri kemičnih reakcijah, kot je sestavljanje makromolekul iz monomerov. Na primer, aminokisline se lahko združijo v peptide le, če so bile predhodno fosforilirane. Mehanski procesi krčenja ali gibanja, prenos topljenca proti koncentracijskemu gradientu in drugi procesi so povezani s porabo energije, shranjene v ATP.
Proces izmenjave energije lahko predstavimo na naslednji način. Visokomolekularne organske snovi v citoplazmi se encimsko, s hidrolizo, pretvorijo v enostavnejše, iz katerih so sestavljene: beljakovine - v aminokisline, poli- in disaharidi - v monosaharide (+ glukoza), maščobe v glicerol in maščobne kisline. Oksidativni procesi so odsotni, sprosti se malo energije, ki se ne porabi in gre v toplotno obliko. Večina celic najprej uporablja ogljikove hidrate. Polisaharidi (škrob v rastlinah in glikogen pri živalih) se hidrolizirajo v glukozo. Oksidacija glukoze poteka v treh fazah: glikoliza, oksidativna dekarboksilacija (Krebsov cikel - cikel citronske kisline) in oksidativna fosforilacija (dihalna veriga). V citoplazmi poteka glikoliza, zaradi katere se ena molekula glukoze razdeli na dve molekuli pirovične kisline s sproščanjem dveh molekul ATP. V odsotnosti kisika se piruvična kislina pretvori v etanol (fermentacija) ali mlečno kislino (anaerobno dihanje).
Če se glikoliza izvaja v živalskih celicah, se molekula glukoze s šestimi ogljiki razgradi na dve molekuli mlečne kisline. Ta proces je večstopenjski. Zaporedoma ga izvaja 13 encimov. Med alkoholno fermentacijo iz molekule glukoze nastaneta dve molekuli etanola in dve molekuli CO2.
Glikoliza je faza, ki je skupna anaerobnemu in aerobnemu dihanju, drugi dve se izvajata le v aerobnih pogojih. Proces oksidacije brez kisika, pri katerem se sprosti in uporabi le del energije metabolitov, je končni proces za anaerobne organizme. V prisotnosti kisika piruvična kislina prehaja v mitohondrije, kjer se zaradi številnih zaporednih reakcij popolnoma aerobno oksidira v H2O in CO2 ob hkratni fosforilaciji ADP v ATP. Hkrati glikoliza daje dve molekuli ATP, dve - Krebsov cikel, 34 - dihalno verigo. Neto donos iz popolne oksidacije ene molekule glukoze v H2O in CO2 je 38 molekul.
Tako se pri aerobnih organizmih končna razgradnja organskih snovi izvede z oksidacijo z atmosferskim kisikom do preprostih anorganskih snovi: CO2 in H2O. Ta proces poteka na kristah mitohondrijev. V tem primeru se sprosti največja količina proste energije, katere pomemben del je rezerviran v molekulah ATP. Zlahka je videti, da aerobna oksidacija celici v največji meri zagotavlja brezplačno energijo.
Zaradi katabolizma se v celici kopičijo z energijo bogate molekule ATP, CO2 in odvečna voda pa se sprostita v zunanje okolje.
Molekule sladkorja, ki niso potrebne za dihanje, se lahko shranijo v celici. Odvečni lipidi se bodisi cepijo, nato pa njihovi produkti cepitve vstopijo v mitohondrije kot substrat za dihanje, ali pa se odložijo v rezervi v citoplazmi v obliki maščobnih kapljic. Beljakovine so zgrajene iz aminokislin, ki vstopajo v celico. Sinteza beljakovin poteka v organelah, imenovanih ribosomi. Vsak ribosom je sestavljen iz dveh poddelcev – velikega in majhnega: oba poddelca vključujeta beljakovinske molekule in molekule RNA.
Ribosomi so pogosto pritrjeni na poseben sistem membran, sestavljen iz rezervoarjev in veziklov, na tako imenovani endoplazmatski retikulum (ER); v celicah, ki proizvajajo veliko beljakovin, je endoplazmatski retikulum pogosto zelo dobro razvit in poln ribosomov. Nekateri encimi so učinkoviti le, če so pritrjeni na membrano. Tu se nahaja večina encimov, ki sodelujejo pri sintezi lipidov. Tako je endoplazmatski retikulum tako rekoč nekakšno namizje celic.
Poleg tega ER razdeli citoplazmo na ločene odseke ali predelke, to je, ločuje različne kemične procese, ki se hkrati pojavljajo v citoplazmi, in s tem zmanjša verjetnost, da bodo ti procesi medsebojno motili.
Pogosto se izdelki, ki jih tvori dana celica, uporabljajo zunaj celice. V takih primerih proteini, sintetizirani na ribosomih, prehajajo skozi membrane endoplazmatskega retikuluma in se pakirajo v membranske vezikle, ki se tvorijo okoli njih, ki se nato izvlečejo iz ER. Ti mehurčki, ki se sploščijo in zložijo drug na drugega, kot palačinke na kupu, tvorijo značilno strukturo, imenovano Golgijev kompleks ali Golgijev aparat. Med bivanjem v Golgijevem aparatu so beljakovine podvržene določenim spremembam. Ko je čas, da zapustijo celico, se membranski mehurčki zlijejo s celično membrano in se izpraznijo ter izlijejo svojo vsebino navzven, torej pride do izločanja z eksocitozo.
Lizosomi nastajajo tudi v Golgijevem aparatu – membranskih vrečkah, ki vsebujejo prebavne encime. Razumevanje, kako celica proizvaja, pakira in izvaža določene beljakovine in kako »ve«, katere beljakovine naj obdrži zase, je ena najbolj fascinantnih vej sodobne citologije.
Membrane katere koli celice se nenehno premikajo in spreminjajo. ER membrane se počasi premikajo po celici. Ločeni deli teh membran se ločijo in tvorijo vezikle, ki začasno postanejo del Golgijevega aparata, nato pa se v procesu eksocitoze združijo s celično membrano.
Kasneje se membranski material vrne v citoplazmo, kjer se ponovno uporabi.
Izmenjava snovi, ki vstopajo v celico ali se iz nje sproščajo navzven, ter izmenjava različnih signalov z mikro- in makrookoljem, poteka skozi zunanjo membrano celice. Kot je znano, je celična membrana lipidni dvosloj, v katerega so vgrajene različne beljakovinske molekule, ki delujejo kot specializirani receptorji, ionski kanali, naprave, ki aktivno prenašajo ali odstranjujejo različne kemikalije, medcelične stike itd. V zdravih evkariontskih celicah so fosfolipidi porazdeljeni v membrana asimetrično: zunanja površina je sestavljena iz sfingomielina in fosfatidilholina, notranja pa iz fosfatidilserina in fosfatidiletanolamina. Ohranjanje takšne asimetrije zahteva porabo energije. Zato je v primeru poškodbe celice, njene okužbe, energetskega stradanja zunanja površina membrane obogatena z zanjo nenavadnimi fosfolipidi, ki postanejo signal drugim celicam in encimom o poškodbi celice z ustrezno reakcijo nanjo. Najpomembnejšo vlogo ima topna oblika fosfolipaze A2, ki razgrajuje arahidonsko kislino in iz zgornjih fosfolipidov tvori lizoforme. Arahidonska kislina je omejujoča povezava za nastanek takšnih vnetnih mediatorjev, kot so eikozanoidi, na lizoforme v membrani pa se vežejo zaščitne molekule - pentraksini (C-reaktivni protein (CRP), prekurzorji amiloidnih proteinov), čemur sledi aktivacija sistem komplementa po klasični poti in uničenje celic.
Struktura membrane prispeva k ohranjanju značilnosti notranjega okolja celice, njenih razlik od zunanjega okolja. To zagotavlja selektivna prepustnost celične membrane, obstoj v njej aktivnih transportnih mehanizmov. Njihova kršitev zaradi neposredne poškodbe, na primer s tetrodotoksinom, ouabainom, tetraetilamonijem ali v primeru nezadostne oskrbe z energijo ustreznih "črpalk", vodi do kršitve sestave elektrolitov celice, spremembe v njeni presnovi. , kršitev specifičnih funkcij - krčenje, prevajanje impulza vzbujanja itd. Kršitev celičnih ionskih kanalov (kalcij, natrij, kalij in klorid) pri človeku je lahko tudi genetsko določena z mutacijo genov, ki so odgovorni za strukturo teh kanalov. Tako imenovane kanalopatije so vzrok za dedne bolezni živčnega, mišičnega in prebavnega sistema. Prekomerni vnos vode v celico lahko privede do njenega razpoka - citolize - zaradi perforacije membrane med aktivacijo komplementa ali napada citotoksičnih limfocitov in naravnih morilcev.
V celično membrano je vgrajenih veliko receptorjev – struktur, ki v kombinaciji z ustreznimi specifičnimi signalnimi molekulami (ligandi) posredujejo signal v celico. To se dogaja prek različnih regulacijskih kaskad, sestavljenih iz encimsko aktivnih molekul, ki se zaporedno aktivirajo in na koncu prispevajo k izvajanju različnih celičnih programov, kot so rast in proliferacija, diferenciacija, gibljivost, staranje in celična smrt. Regulatorne kaskade so precej številne, vendar njihovo število še ni v celoti določeno. Sistem receptorjev in z njimi povezane regulacijske kaskade obstajajo tudi znotraj celice; ustvarjajo določeno regulativno mrežo s točkami koncentracije, porazdelitve in izbire nadaljnje signalne poti glede na funkcionalno stanje celice, stopnjo njenega razvoja in hkratno delovanje signalov drugih receptorjev. Rezultat tega je lahko zaviranje ali ojačanje signala, njegova smer vzdolž drugačne regulacijske poti. Tako receptorski aparat kot poti signalne transdukcije skozi regulacijske kaskade, na primer do jedra, so lahko motene zaradi genetske okvare, ki se pojavi kot prirojena napaka na ravni organizma ali zaradi somatske mutacije v določeni celici. tip. Te mehanizme lahko poškodujejo povzročitelji infekcij, toksini, spremenijo pa se tudi med staranjem. Končna faza tega je lahko kršitev funkcij celice, procesov njene proliferacije in diferenciacije.
Molekule, ki igrajo pomembno vlogo v procesih medcelične interakcije, se nahajajo tudi na površini celic. Ti lahko vključujejo celične adhezijske proteine, antigene tkivne združljivosti, tkivno specifične, diferenciacijske antigene itd. Spremembe v sestavi teh molekul povzročijo kršitev medceličnih interakcij in lahko povzročijo aktivacijo ustreznih mehanizmov za eliminacijo takšnih celic, ker predstavljajo določeno nevarnost za celovitost telesa kot rezervoar okužbe, zlasti virusne, ali kot potencialni iniciator rasti tumorja.
Kršitev oskrbe celice z energijo
Vir energije v celici je hrana, po razgradnji katere se energija sprosti do končnih snovi. Mitohondriji so glavno mesto proizvodnje energije, v kateri se snovi oksidirajo s pomočjo encimov dihalne verige. Oksidacija je glavni dobavitelj energije, saj se zaradi glikolize iz enake količine oksidacijskih substratov (glukoze) ne sprosti več kot 5 % energije v primerjavi z oksidacijo. Približno 60 % energije, ki se sprosti med oksidacijo, se akumulira z oksidativno fosforilacijo v makroergičnih fosfatih (ATP, kreatin fosfat), preostanek se razprši kot toplota. V prihodnosti bo celica uporabljala visokoenergijske fosfate za procese, kot so črpanje, sinteza, delitev, gibanje, izločanje itd. Obstajajo trije mehanizmi, katerih poškodba lahko povzroči motnje v oskrbi celice z energijo. : prvi je mehanizem za sintezo encimov energetske presnove, drugi je mehanizem oksidativne fosforilacije, tretji mehanizem porabe energije.
Kršitev transporta elektronov v dihalni verigi mitohondrijev ali odklop oksidacije in fosforilacije ADP z izgubo protonskega potenciala - gonilne sile nastajanja ATP, vodi v oslabitev oksidativne fosforilacije na način, da se večina energije razprši v oblika toplote in število makroergičnih spojin se zmanjša. Prekinitev oksidacije in fosforilacije pod vplivom adrenalina uporabljajo celice homoiotermnih organizmov za povečanje proizvodnje toplote ob ohranjanju konstantne telesne temperature med ohlajanjem ali njenega povečanja med vročino. Pri tirotoksikozi opazimo pomembne spremembe v strukturi mitohondrijev in energijskem metabolizmu. Te spremembe so sprva reverzibilne, po določenem času pa postanejo nepopravljive: mitohondriji se razpadejo ali nabreknejo, izgubijo kriste, se spremenijo v vakuole in sčasoma kopičijo snovi, kot so hialin, feritin, kalcij, lipofuscin. Pri bolnikih s skorbutom se mitohondriji zlijejo v hondriosfere, verjetno zaradi poškodbe membrane s peroksidnimi spojinami. Pomembne poškodbe mitohondrijev nastanejo pod vplivom ionizirajočega sevanja, pri preoblikovanju normalne celice v maligno.
Mitohondriji so močan depo kalcijevih ionov, kjer je njegova koncentracija za nekaj redov višja od koncentracije v citoplazmi. Ko so mitohondriji poškodovani, kalcij vstopi v citoplazmo, kar povzroči aktivacijo proteinaz s poškodbo znotrajceličnih struktur in motnjo funkcij ustrezne celice, na primer kalcijeve kontrakture ali celo "kalcijevo smrt" v nevronih. Zaradi kršitve funkcionalne sposobnosti mitohondrijev se močno poveča tvorba peroksidnih spojin prostih radikalov, ki imajo zelo visoko reaktivnost in zato poškodujejo pomembne celične komponente - nukleinske kisline, beljakovine in lipide. Ta pojav opazimo med tako imenovanim oksidativnim stresom in ima lahko negativne posledice za obstoj celice. Tako poškodbo zunanje mitohondrijske membrane spremlja sproščanje v citoplazmo snovi, ki jih vsebuje medmembranski prostor, predvsem citokroma C in nekaterih drugih biološko aktivnih snovi, ki sprožijo verižne reakcije, ki povzročajo programirano celično smrt – apoptozo. S poškodbo mitohondrijske DNK reakcije prostih radikalov izkrivljajo genetske informacije, potrebne za tvorbo določenih encimov dihalne verige, ki se proizvajajo posebej v mitohondrijih. To vodi do še večje motnje oksidativnih procesov. Na splošno je intrinzični genetski aparat mitohondrijev v primerjavi z genetskim aparatom jedra manj zaščiten pred škodljivimi vplivi, ki lahko spremenijo genetske informacije, ki so v njem kodirane. Posledično se mitohondrijska disfunkcija pojavlja skozi vse življenje, na primer v procesu staranja, med maligno transformacijo celice, pa tudi v ozadju dednih mitohondijskih bolezni, povezanih z mutacijo mitohondrijske DNK v jajčecu. Trenutno je opisanih več kot 50 mitohondrijskih mutacij, ki povzročajo dedne degenerativne bolezni živčnega in mišičnega sistema. Na otroka se prenašajo izključno od matere, saj mitohondriji semenčic niso del zigote in s tem novega organizma.
Kršitev ohranjanja in prenosa genetskih informacij
Celično jedro vsebuje večino genetskih informacij in tako zagotavlja njegovo normalno delovanje. S pomočjo selektivne genske ekspresije usklajuje delo celice v interfazi, shranjuje genetske informacije, poustvarja in prenaša genetski material v procesu celične delitve. Replikacija DNK in transkripcija RNA potekata v jedru. Različni patogeni dejavniki, kot so ultravijolično in ionizirajoče sevanje, oksidacija prostih radikalov, kemikalije, virusi, lahko poškodujejo DNK. Ocenjuje se, da vsaka celica toplokrvne živali v 1 dnevu. izgubi več kot 10.000 baz. Temu je treba dodati še kršitve pri kopiranju med delitvijo. Če bi ta poškodba vztrajala, celica ne bi mogla preživeti. Zaščita je v obstoju močnih popravljalnih sistemov, kot so ultravijolična endonukleaza, reparativna replikacija in rekombinacijski sistemi popravljanja, ki nadomestijo poškodbe DNK. Genetske okvare v reparativnih sistemih povzročajo razvoj bolezni zaradi povečane občutljivosti na dejavnike, ki poškodujejo DNK. To je pigmentirana kseroderma, pa tudi nekateri sindromi pospešenega staranja, ki jih spremlja povečana nagnjenost k nastanku malignih tumorjev.
Sistem regulacije procesov replikacije DNK, transkripcije sporočilne RNA (mRNA), prevajanja genetskih informacij iz nukleinskih kislin v strukturo beljakovin je precej zapleten in večstopenjski. Poleg regulatornih kaskad, ki sprožijo delovanje več kot 3000 transkripcijskih faktorjev, ki aktivirajo določene gene, obstaja tudi večstopenjski regulativni sistem, ki ga posredujejo majhne molekule RNA (interferirajoče RNA; RNAi). Človeški genom, ki je sestavljen iz približno 3 milijard purinskih in pirimidinskih baz, vsebuje le 2 % strukturnih genov, odgovornih za sintezo beljakovin. Ostale zagotavljajo sintezo regulatornih RNA, ki skupaj s transkripcijskimi faktorji aktivirajo ali blokirajo delo strukturnih genov na ravni DNK v kromosomih ali vplivajo na procese prevajanja sporočilne RNA (mRNA) med tvorbo polipeptidne molekule v kromosomih. citoplazmo. Kršitev genetskih informacij se lahko pojavi tako na ravni strukturnih genov kot na regulativnem delu DNK z ustreznimi manifestacijami v obliki različnih dednih bolezni.
V zadnjem času se veliko pozornosti posveča spremembam v genskem materialu, ki nastanejo med individualnim razvojem organizma in so povezane z inhibicijo ali aktivacijo določenih delov DNK in kromosomov zaradi njihove metilacije, acetilacije in fosforilacije. Te spremembe trajajo dolgo časa, včasih skozi celotno življenje organizma od embriogeneze do starosti in se imenujejo epigenomsko dedovanje.
Razmnoževanje celic s spremenjeno genetsko informacijo preprečujejo tudi sistemi (faktorji) nadzora mitotičnega cikla. Vzajemno delujejo s ciklin odvisnimi protein kinazami in njihovimi katalitičnimi podenotami - ciklini - in blokirajo prehod celotnega mitotičnega cikla v celici, ustavi delitev na meji med predsintetično in sintetično fazo (blok G1 / S), dokler se popravilo DNK ne zaključi. , in če je to nemogoče, sprožijo programirane celice smrti. Ti dejavniki vključujejo gen p53, katerega mutacija povzroči izgubo nadzora nad proliferacijo transformiranih celic; pojavlja se pri skoraj 50 % rakavih obolenj pri ljudeh. Druga kontrolna točka prehoda mitotičnega cikla se nahaja na meji G2/M. Tukaj se pravilno porazdelitev kromosomskega materiala med hčerinskimi celicami v mitozi ali mejozi nadzoruje s pomočjo kompleksa mehanizmov, ki nadzorujejo celično vreteno, središče in centromere (kinetohore). Neučinkovitost teh mehanizmov vodi do kršitve porazdelitve kromosomov ali njihovih delov, kar se kaže v odsotnosti katerega koli kromosoma v eni od hčerinskih celic (aneuploidija), prisotnosti dodatnega kromosoma (poliploidija), ločitvi del kromosoma (delecija) in njegov prenos na drug kromosom (translokacija). Takšne procese zelo pogosto opazimo med razmnoževanjem maligno degeneriranih in transformiranih celic. Če se to zgodi med mejozo z zarodnimi celicami, vodi bodisi do smrti ploda v zgodnji fazi embrionalnega razvoja bodisi do rojstva organizma s kromosomsko boleznijo.
Nenadzorovana reprodukcija celic med rastjo tumorja nastane kot posledica mutacij v genih, ki nadzorujejo celično proliferacijo in se imenujejo onkogeni. Med več kot 70 trenutno znanimi onkogeni je večina komponent regulacije celične rasti, nekateri so transkripcijski faktorji, ki uravnavajo gensko aktivnost, pa tudi dejavniki, ki zavirajo delitev in rast celic. Drug dejavnik, ki omejuje prekomerno širjenje (širjenje) proliferirajočih celic, je skrajšanje koncev kromosomov - telomerov, ki se zaradi čisto sterične interakcije ne morejo v celoti replicirati, zato se po vsaki delitvi celice telomeri skrajšajo za določen del podlag. Tako proliferirajoče celice odraslega organizma po določenem številu delitev (običajno od 20 do 100, odvisno od vrste organizma in njegove starosti) izčrpajo dolžino telomera in nadaljnja replikacija kromosomov se ustavi. Ta pojav se ne pojavlja v spermatogenem epiteliju, enterocitih in embrionalnih celicah zaradi prisotnosti encima telomeraze, ki obnavlja dolžino telomerov po vsaki delitvi. V večini celic odraslih organizmov je telomeraza blokirana, vendar se na žalost aktivira v tumorskih celicah.
Povezava med jedrom in citoplazmo, transport snovi v obe smeri poteka skozi pore v jedrski membrani s sodelovanjem posebnih transportnih sistemov s porabo energije. Tako se energijske in plastične snovi, signalne molekule (transkripcijski faktorji) transportirajo v jedro. Povratni tok prinaša v citoplazmo molekule mRNA in prenosne RNA (tRNA), ribosome, potrebne za sintezo beljakovin v celici. Enak način transporta snovi je značilen za viruse, zlasti kot je HIV. Svoj genetski material prenesejo v jedro gostiteljske celice z njegovo nadaljnjo vključitvijo v gostiteljski genom in prenosom novonastale virusne RNK v citoplazmo za nadaljnjo sintezo beljakovin novih virusnih delcev.
Kršitev sinteznih procesov
Procesi sinteze beljakovin potekajo v cisternah endoplazmatskega retikuluma, tesno povezanih s porami v jedrski membrani, skozi katere ribosomi, tRNA in mRNA vstopajo v endoplazmatski retikulum. Tu se izvaja sinteza polipeptidnih verig, ki kasneje dobijo končno obliko v agranularnem endoplazmatskem retikulumu in lamelarnem kompleksu (Golgijev kompleks), kjer so podvržene post-translacijski modifikaciji in povezavi z molekulami ogljikovih hidratov in lipidov. Novonastale beljakovinske molekule ne ostanejo na mestu sinteze, temveč s pomočjo kompleksnega reguliranega procesa, ki se imenuje protein kineza, se aktivno prenašajo v tisti izolirani del celice, kjer bodo opravljale svojo predvideno funkcijo. V tem primeru je zelo pomemben korak strukturiranje prenesene molekule v ustrezno prostorsko konfiguracijo, ki lahko opravlja svojo lastno funkcijo. Takšno strukturiranje poteka s pomočjo posebnih encimov ali na matriksu specializiranih beljakovinskih molekul - chaperonov, ki pomagajo, da beljakovinska molekula, na novo nastala ali zaradi zunanjega vpliva spremenjena, pridobi pravilno tridimenzionalno strukturo. V primeru škodljivega učinka na celico, ko obstaja možnost kršitve strukture beljakovinskih molekul (na primer s povišanjem telesne temperature, infekcijskim procesom, zastrupitvijo), se koncentracija spremljevalcev v celici močno narašča. Zato se takšne molekule imenujejo tudi stresne beljakovine, oz beljakovine toplotnega šoka. Kršitev strukturiranja proteinske molekule vodi do tvorbe kemično inertnih konglomeratov, ki se odlagajo v celici ali zunaj nje, če bo amiloidoza, Alzheimerjeva bolezen itd. okvarjena. Ta situacija se pojavi pri tako imenovanih prionskih boleznih (scrappie pri ovcah, steklina pri kravah, kuru, Creutzfeldt-Jakobova bolezen pri ljudeh), ko okvara enega od membranskih proteinov živčne celice povzroči naknadno kopičenje inertnih mas v notranjosti. celice in motnje njene vitalne aktivnosti.
Kršitev sinteznih procesov v celici se lahko pojavi na različnih stopnjah: transkripcija RNA v jedru, translacija polipeptidov v ribosomih, posttranslacijska modifikacija, hipermetilacija in glikozilacija bež molekule, transport in porazdelitev beljakovin v celici ter njihova odstranitev. na zunanjost. V tem primeru lahko opazimo povečanje ali zmanjšanje števila ribosomov, razgradnjo poliribosomov, širjenje cistern granularnega endoplazmatskega retikuluma, izgubo ribosomov z njim, nastanek veziklov in vakuol. Torej, v primeru zastrupitve z bledo krastačo se poškoduje encim RNA polimeraza, kar moti transkripcijo. Toksin davice, ki inaktivira faktor raztezanja, moti procese prevajanja in povzroči poškodbe miokarda. Razlog za kršitev sinteze nekaterih specifičnih beljakovinskih molekul so lahko povzročitelji infekcij. Herpesvirusi na primer zavirajo sintezo in izražanje molekul antigena MHC, kar jim omogoča, da se delno izognejo imunskemu nadzoru, bacili kuge pa zavirajo sintezo mediatorjev akutnega vnetja. Pojav nenavadnih beljakovin lahko ustavi njihovo nadaljnjo razgradnjo in povzroči kopičenje inertnega ali celo strupenega materiala. K temu lahko v določeni meri prispeva tudi prekinitev procesov razpadanja.
Kršitev procesov razpadanja
Hkrati s sintezo beljakovin v celici se nenehno pojavlja njen razpad. V normalnih pogojih ima to pomemben regulacijski in formativni pomen, na primer med aktivacijo neaktivnih oblik encimov, beljakovinskih hormonov in beljakovin mitotičnega cikla. Normalna rast in razvoj celic zahtevata natančno nadzorovano ravnovesje med sintezo in razgradnjo beljakovin in organelov. Vendar pa se v procesu sinteze beljakovin zaradi napak pri delovanju aparata za sintezo, nenormalnega strukturiranja beljakovinske molekule, njene poškodbe s kemičnimi in bakterijskimi sredstvi nenehno tvori precej veliko število okvarjenih molekul. Po nekaterih ocenah je njihov delež približno tretjina vseh sintetiziranih beljakovin.
Celice sesalcev imajo več glavnih Poti razgradnje beljakovin: preko lizosomskih proteaz (pentidne hidrolaze), od kalcija odvisnih proteinaz (endopeptidaze) in proteasomskega sistema. Poleg tega obstajajo tudi specializirane proteinaze, kot so kaspaze. Glavna organela, v kateri poteka razgradnja snovi v evkariontskih celicah, je lizosom, ki vsebuje številne hidrolitične encime. Zaradi procesov endocitoze in različnih vrst avtofagije v lizosomih in fagolizosomih se uničijo tako okvarjene beljakovinske molekule kot celotne organele: poškodovani mitohondriji, deli plazemske membrane, nekateri zunajcelični proteini, vsebina sekretornih zrnc.
Pomemben mehanizem razgradnje beljakovin je proteasom, kompleksna multikatalitična proteinazna struktura, lokalizirana v citosolu, jedru, endoplazmatskem retikulumu in na celični membrani. Ta encimski sistem je odgovoren za razgradnjo poškodovanih beljakovin, pa tudi zdravih beljakovin, ki jih je treba odstraniti za normalno delovanje celic. V tem primeru se proteini, ki jih je treba uničiti, predhodno združijo s specifičnim polipeptidom ubikvitina. Vendar pa se lahko beljakovine, ki niso vsepovsodne, delno uničijo tudi v proteasomih. Razgradnja proteinske molekule v proteasomih na kratke polipeptide (obdelava) z njihovo naknadno predstavitvijo skupaj z molekulami MHC tipa I je pomemben člen pri izvajanju imunskega nadzora antigenske homeostaze telesa. Ko je funkcija proteasoma oslabljena, pride do kopičenja poškodovanih in nepotrebnih beljakovin, kar spremlja staranje celic. Kršitev razgradnje ciklin odvisnih beljakovin vodi do kršitve delitve celic, razgradnje sekretornih beljakovin - do razvoja cistofibroze. Nasprotno pa povečanje funkcije proteasoma spremlja izčrpavanje telesa (AIDS, rak).
Z genetsko določenimi kršitvami razgradnje beljakovin organizem ni sposoben preživetja in umre v zgodnjih fazah embriogeneze. Če je razgradnja maščob ali ogljikovih hidratov motena, se pojavijo akumulacijske bolezni (tezavrizme). Hkrati se znotraj celice nabira prevelika količina določenih snovi oziroma produktov njihove nepopolne razgradnje – lipidov, polisaharidov, kar bistveno poškoduje delovanje celice. Najpogosteje ga opazimo v jetrnih epiteliocitih (hepatocitih), nevronih, fibroblastih in makrofagocitih.
Pridobljene motnje v procesih razpadanja snovi se lahko pojavijo kot posledica patoloških procesov (na primer distrofija beljakovin, maščob, ogljikovih hidratov in pigmentov) in jih spremlja tvorba nenavadnih snovi. Kršitve v sistemu lizosomske proteolize vodijo do zmanjšanja prilagajanja med stradanjem ali povečano obremenitvijo, do pojava nekaterih endokrinih disfunkcij - zmanjšanja ravni insulina, tiroglobulina, citokinov in njihovih receptorjev. Kršitve razgradnje beljakovin upočasnijo hitrost celjenja ran, povzročijo razvoj ateroskleroze in vplivajo na imunski odziv. Pri hipoksiji, spremembah znotrajceličnega pH, sevalnih poškodbah, za katere je značilna povečana peroksidacija membranskih lipidov, pa tudi pod vplivom lizosomotropnih snovi - bakterijskih endotoksinov, presnovkov strupenih gliv (sporofusarin), kristalov silicijevega oksida - stabilnost lizosomske membrane sprememb, aktivirani lizosomski encimi se sproščajo v citoplazmo, kar povzroči uničenje celičnih struktur in njeno smrt.
Poglavje 1
OSNOVE CELIČNE FIZIOLOGIJE
I. Dudel
plazemska membrana . Živalske celice so omejene s plazemsko membrano (slika 1.1). Podrobneje se bomo zadržali na njeni zgradbi, ki je zelo podobna zgradbi številnih znotrajceličnih membran. Glavna matrika membrane je sestavljena iz lipidov predvsem fosfatidilholin. Ti lipidi so sestavljeni iz hidrofilne glavne skupine, na katero so vezane dolge hidrofobne ogljikovodične verige. V vodi takšni lipidi spontano tvorijo dvoslojni film debeline 4–5 nm, v katerem so hidrofilne skupine obrnjene proti vodnemu mediju, hidrofobne ogljikovodične verige pa so razporejene v dve vrsti in tvorijo brezvodno lipidno fazo. Celične membrane so lipidni dvosloji te vrste in vsebujejo glikolipide, holesterol in fosfolipide (slika 1.2). Hidrofilni del glikolipidov tvorijo oligosaharidi. Glikolipidi se vedno nahajajo na zunanji površini plazemske membrane, oligosaharidni del molekule pa je usmerjen kot las, potopljen v okolje. Razpršene med fosfolipidi v skoraj enakih količinah molekul holesterola stabilizirajo membrano. Porazdelitev različnih lipidov v notranjih in zunanjih plasteh membrane ni enaka in tudi znotraj iste plasti so območja, v katerih so koncentrirane določene vrste lipidov. Tako neenakomerna porazdelitev
riž. 1.1. Shematska risba celice, ki prikazuje najpomembnejše organele
verjetno ima nek, še nejasen funkcionalni pomen.
Glavni funkcionalni elementi, potopljeni v relativno inertno lipidno matriko membrane, so veverice(slika 1.2). Masni delež beljakovin se v različnih membranah giblje od 25 do 75 %, a ker so beljakovinske molekule veliko večje od lipidnih molekul, je 50 masnih % enakovredno razmerju 1 beljakovinska molekula proti 50 molekulam lipidov. Nekateri proteini prodrejo skozi membrano od njene zunanje do notranje površine, drugi pa so fiksirani v eni plasti. Proteinske molekule so običajno orientirane tako, da so njihove hidrofobne skupine potopljene v lipidno membrano, polarne hidrofilne skupine na površini membrane pa so potopljene v vodno fazo. Mnogi proteini zunanje površinske membrane so glikoproteini; njihove hidrofilne saharidne skupine se soočajo z zunajceličnim okoljem.
Membranski sistem znotrajceličnih organelov .
Približno polovico volumna celice zasedajo organele, izolirane iz citosola z membranami. Celotna površina membran znotrajceličnih organelov je vsaj 10-krat večja od površine plazemske membrane. Najbolj razširjen membranski sistem je Endoplazemski retikulum, ki predstavljajo omrežje

riž. 1.2.Shematski prikaz plazemske membrane. Beljakovine so vgrajene v fosfolipidni dvosloj, pri čemer nekatere pokrivajo dvosloj, medtem ko so druge zasidrane le na zunanjo ali notranjo plast
močno zvite tubule ali vrečaste podolgovate strukture; velika območja endoplazmatskega retikuluma so posejana z ribosomi; tak retikulum imenujemo zrnat ali hrapav (slika 1.1). golgijev aparat sestoji tudi iz membransko vezanih lamel, od katerih se odcepijo mehurčki ali mehurčki (slika 1.1). Lizosomi in peroksisomi so majhni specializirani mehurčki. V vseh teh raznolikih organelah vsebujeta membrana in prostor, ki ga pokrivata, posebne sklope encimov; znotraj organelov se kopičijo posebni presnovni produkti, ki se uporabljajo za izvajanje različnih funkcij organelov.
Jedroin mitohondrije razlikujejo po tem, da je vsaka od teh organelov obdana z dvema membranama. Jedro je odgovorno za kinetični nadzor presnove; nagubana notranja mitohondrijska membrana je mesto oksidativnega metabolizma; tu se zaradi oksidacije piruvata ali maščobnih kislin sintetizira visokoenergijska spojina adenozin trifosfat (ATP ali ATP).
citoskelet . Citoplazme, ki obdaja organele, nikakor ne moremo šteti za amorfno; prežeta ga mreža citoskeleta. Citoskelet je sestavljen iz mikrotubul, aktinskih filamentov in vmesnih filamentov (slika 1.1). mikrotubule imajo zunanji premer približno 25 nm; nastanejo kot običajni polimer kot posledica sestavljanja proteinskih molekul tubulina. aktinski filamenti- kontraktilna vlakna, ki se nahajajo v sloju blizu membrane in po celi celici - večinoma sodelujejo v procesih, povezanih z gibanjem. Vmesni filamenti sestavljeni iz blokov različne kemične sestave v različnih vrstah celic; tvorijo različne povezave med drugim dvema elementoma zgoraj omenjenega citoskeleta. S citoskeletom so povezane tudi organele in plazemska membrana, ki ne ohranja le oblike celice in položaja organelov v njej, temveč določa tudi spremembo oblike celice in njeno gibljivost.
Cytosol . Približno polovico volumna celice zaseda citosol. Ker je približno 20% (po masi) beljakovin, je bolj gel kot vodna raztopina. Majhne molekule, vključno z organskimi in anorganskimi ioni, raztopljen v vodni fazi. Med celico in okoljem (zunajcelični prostor) poteka izmenjava ionov; ti procesi izmenjave bodo obravnavani v naslednjem razdelku. Koncentracija ionov v zunajceličnem prostoru se vzdržuje s precejšnjo natančnostjo na konstantni ravni; znotrajcelična koncentracija vsakega od ionov ima tudi specifično raven, ki se razlikuje od tiste zunaj celice (tabela 1.1). Najpogostejši kation v zunajceličnem okolju je Na+ v celici je njegova koncentracija več kot 10-krat nižja. Nasprotno, znotraj celice je koncentracija K + najvišja, zunaj celice je nižja za več kot red velikosti. Največji gradient med zunajcelično in znotrajcelično koncentracijo obstaja za Ca 2+, katerega koncentracija prostih ionov znotraj celice je vsaj 10.000-krat nižja kot zunaj nje. Vsi ioni niso raztopljeni v citosolu; nekateri od njih se adsorbirajo na beljakovine ali odložijo v organele. Na primer, v primeru Ca 2+ je vezanih ionov veliko več kot prostih. Večina citosolnih beljakovin je encimov, s sodelovanjem katerih se izvajajo številni procesi vmesne presnove: glikoliza in glukoneogeneza, sinteza ali uničenje aminokislin, sinteza beljakovin na ribosomih (slika 1.1). Citosol vsebuje tudi lipidne kapljice in glikogenska zrnca, ki služijo kot rezerve pomembnih molekul.
Tabela 1.1.Intra- in zunajcelične koncentracije ionov v mišičnih celicah homoiotermnih živali. AMPAK – – “celični anioni z visoko molekulsko maso”
|
Intracelularna koncentracija |
Zunajcelična koncentracija |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
Z l- |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO 3 - |
8 mmol/l |
HCO 3 - |
27 mmol/l |
|
A- |
155 mmol/l |
Drugo kationov |
5 mmol/l |
|
Potencial mirovanja -90 mV |
|||
1.2. Izmenjava snovi med celico in okoljem
Na kratko smo opisali strukturo celice, da bi lahko s tem opisom pregledali osnove celične fiziologije. Celice v nobenem primeru ne moremo šteti za statično tvorbo, saj poteka stalna izmenjava snovi med različnimi znotrajceličnimi oddelki, pa tudi med oddelki in okoljem. Strukture celice so v dinamičnem ravnovesju, interakcija celic med seboj in z zunanjim okoljem pa je nujen pogoj za ohranjanje življenja delujočega organizma. V tem poglavju bomo obravnavali temeljne mehanizme takšne izmenjave. V kasnejših poglavjih bodo ti mehanizmi obravnavani v povezavi z živčno celico in njenimi funkcijami;
vendar so enaki mehanizmi osnova delovanja vseh drugih organov.
Difuzija.Najpreprostejši postopek premikanja snovi je difuzija. V raztopinah (ali plinih) se atomi in molekule prosto gibljejo, razlika v koncentracijah pa je uravnotežena z difuzijo. Razmislite o dveh prostorninah, napolnjenih s tekočino ali plinom (slika 1.3), v katerih imajo snovi koncentracije c1 in c2 in ločena s plastjo s površino A in debelino d. Pretok snovi m v času t opisano Fickov prvi zakon difuzije:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
kjer je D difuzijski koeficient, ki je konstanten za dano snov, topilo in temperaturo. V bolj splošni obliki, za razliko koncentracije dc na razdalji dx
dm/dt= -D A dc/dx,(2)
pretok skozi odsek A je sorazmeren z gradientom koncentracije dc/dx . Predznak minus se pojavi v enačbi, ker je sprememba koncentracije v smeri x negativna.
Difuzija je najpomembnejši proces, s katerim se večina molekul v vodnih raztopinah premika na kratke razdalje. To velja tudi za njihovo gibanje v celici, v kolikor difuzije ne ovirajo membrane. Številne snovi lahko prosto difundirajo skozi lipidne membrane, zlasti voda in raztopljeni plini, kot sta O 2 in CO 2 . topen v maščobi

riž. 1.3.Kvantitativna shema difuzije. Oba prostora sta ločena s plastjo debelinedin območje AMPAK. C; - visoka koncentracija delcev v levem delu volumna, C:, - nizka koncentracija delcev v desnem deli, rožnata površina je koncentracijski gradient v difuzijskem sloju. Difuzijski tok dm/dt – glej enačba (1)
snovi tudi dobro difundirajo skozi membrane; to velja tudi za relativno majhne polarne molekule, kot sta etanol in sečnina, medtem ko sladkorji s težavo prehajajo skozi lipidno plast. Hkrati so lipidne plasti praktično neprepustne za nabite molekule, tudi za anorganske ione. Za neelektrolite se difuzijska enačba (1) običajno pretvori s kombiniranjem značilnosti membrane in difuzijske snovi v eno parameter-prepustnost (P):
dm/dt=P AD c.(3)
Na sl. 1,4 v primerjavi prepustnost (P) lipidne membrane za različne molekule.
Difuzija skozi membranske pore . Plazemska membrana (in druge celične membrane) so prepustne ne le za snovi, ki difundirajo skozi lipidno plast, ampak tudi za številne ione, sladkorje, aminokisline in nukleotide. Te snovi prečkajo membrano skozi pore, ki jih tvorijo transportne beljakovine, vgrajena v membrano. Znotraj takšnih beljakovin je z vodo napolnjen kanal s premerom manj kot 1 nm, skozi katerega lahko difundirajo majhne molekule. Premikajo se po koncentracijskem gradientu in če nosijo naboj, potem njihovo gibanje skozi kanale uravnava tudi membranski potencial. Membranski kanali so relativno selektivni

riž. 1.4.Prepustnost umetnih lipidnih dvoplastov za različne snovi
glede na vrsto molekul, ki lahko prehajajo skozi njih, obstajajo na primer kalijevi, natrijevi in kalcijevi kanali, od katerih je vsak neprepusten za skoraj vse ione razen določenega. Takšne selektivnost zaradi naboja ali strukture veznih mest v stenah kanala, kar olajša transport določene molekule in preprečuje prodiranje drugih snovi skozi kanal (sl. 1,5, A) .
Za vedenjem membranski ionski kanali To je enostavno opazovati, saj je mogoče izmeriti tok, ki nastane med gibanjem ionov, in to celo za en sam kanal. Dokazano je, da kanali spontano in z visoko frekvenco spreminjajo svoje stanje iz odprtega v zaprto. Za kalijev kanal so značilni tokovni impulzi z amplitudo približno 2 pA (2 10 -12 A) in trajanjem nekaj milisekund (glej sliko 2.12, str. 37) [3]. V tem obdobju skozenj preide več deset tisoč ionov. Prehod proteinov iz ene konformacije v drugo preučujemo z rentgensko difrakcijo, Mössbauerjevo spektroskopijo in jedrsko magnetno resonanco (NMR). Beljakovine so tako zelo dinamične, mobilne strukture in kanal skozi protein ni le toga, z vodo napolnjena cev (slika 1.5A), ampak labirint hitro premikajočih se molekularnih skupin in nabojev. Ta dinamični odziv kanala se odraža v energijski profil kanala, prikazano na sl. 1.5, B. Tukaj abscisa prikazuje dolžino kanala od zunanje raztopine s koncentracijo ionov C 0 in potencialom 0 do notranje raztopine s koncentracijo C 1 in potencialom E. Os y

riž. 1.5.A. Shema proteina, ki tvori kalijev kanal, vgrajen v lipidni dvosloj plazemske membrane. Na "steno" kanala so pritrjeni štirje negativni naboji. B. Shematski energijski profil kanala, prikazan na sl. A. Na osi y so narisane vrednosti kinetične energije, potrebne za prehod kanala; vzdolž osi abscise, razdalje med notranjo in zunanjo površino membrane. Energetski minimumi ustrezajo veznim mestom pozitivno nabitih ionov s fiksnimi negativnimi naboji v steni kanala. Energijski maksimumi ustrezajo difuzijskim oviram v kanalu. Domneva se, da konformacija proteina kanala spontano niha; možnosti za energijske profile so prikazane s polnimi in črtkanimi črtami; ta nihanja močno olajšajo vezavo ionov pri premagovanju energijske pregrade (vendar s spremembami)
prikazane so energijske ravni iona na mestih vezave kanalov; vrh v grafu predstavlja prepustnostno pregrado, ki jo mora ionska energija premagati, da bi prodrla v kanal, “dip” grafa pa predstavlja relativno stabilno stanje (vezavo). Kljub obstrukciji energijskega vrha lahko ion prodre v kanal, če se energijski profil spontano cikli; ion se lahko tako nenadoma pojavi "na drugi strani" energijskega vrha in se lahko še naprej premika v celico. Glede na naboj, velikost in stopnjo hidracije iona ter njegove sposobnosti vezave na strukture sten kanalov se energijski profil kanala za različne ione razlikuje, kar lahko pojasni selektivnost posameznih vrst kanalov.
Difuzijsko ravnovesje ionov . Difuzija različnih ionov skozi membranske kanale naj vodi k odpravi razlik v koncentracijah med zunaj- in znotrajceličnim okoljem. Vendar, kot je razvidno iz tabele. 1.1, takšne razlike še vedno obstajajo, zato jih mora biti nekaj ravnotežje med difuzijo in drugimi transportnimi procesi čez membrano. Naslednja dva razdelka obravnavata načine, na katere se vzpostavi takšno ravnotežje. V primeru ionov na difuzijsko ravnotežje vpliva njihov naboj. Difuzijo nenabitih molekul zagotavlja razlika v koncentraciji dc , in ko se koncentracije izenačijo, se dejanski transport ustavi. Na nabite delce dodatno vpliva električno polje. Na primer, ko kalijev ion izstopi iz celice vzdolž njegovega koncentracijskega gradienta, nosi en pozitiven naboj. Tako znotrajcelično okolje postane bolj negativno nabito, kar ima za posledico potencialno razliko čez membrano. Znotrajcelični negativni naboj preprečuje, da bi novi kalijevi ioni zapustili celico, tisti ioni, ki kljub temu zapustijo celico, pa bodo še povečali naboj na membrani. Pretok kalijevih ionov se ustavi, ko delovanje električnega polja kompenzira difuzijski tlak zaradi razlike v koncentracijah. Ioni še naprej prehajajo skozi membrano, vendar v enakih količinah v obe smeri. Zato za dano razliko v koncentracijah ionov na membrani obstaja ravnotežni potencial E ion pri katerem se pretok ionov skozi membrano ustavi. Ravnotežni potencial je mogoče enostavno določiti z uporabo Nernstove enačbe:
Eion= RT/ zF* lnven/ C v(4)
kjer je R je plinska konstanta, T je absolutna temperatura, z je valenca ionov (negativna za anione) ven je koncentracija zunajceličnih ionov, C v je znotrajcelična koncentracija iona, F Faradayeva številka. Če v enačbo nadomestimo konstante, potem je pri telesni temperaturi (T = 310 K) ravnotežni potencial za kalijeve ione E K je enako:
Ek= –61 mB dnevnik /(5)
Če [ K + izhod ]/[ K + vhod ] = 39, kot sledi iz tabele. 1.1 torej
Ek= -61 m B log 39= -97 mV.
Dejansko je bilo ugotovljeno, da imajo vse celice membranski potencial; v mišičnih celicah sesalcev je njegova raven približno -90 mV. Glede na pogoje in relativne koncentracije ionov imajo lahko celice membranski potencial v razponu od -40 do -120 mV. Za celico v zgornjem primeru (tabela 1.1) možnost počitka, enako približno -90 mV, kaže, da so tokovi kalijevih ionov skozi membranske kanale približno v ravnotežju. To ni presenetljivo, saj je najverjetneje odprto stanje kalijevih kanalčkov v mirni membrani; membrana je najbolj prepustna za kalijeve ione. Membranski potencial pa določajo tudi tokovi drugih ionov.
Enostavnost, s katero lahko nenabiti delci difundirajo skozi membrano, je kvantificirana v enačbi (3). Prepustnost za nabite delce opisano z nekoliko bolj zapleteno enačbo:
P= m RT/ dF(6)
kje mje mobilnost iona v membrani, d - debelina membrane, a R, T in F znane termodinamične konstante. Tako določene vrednosti prepustnosti za različne ione lahko uporabimo za izračun membranskega potenciala Em ko kalijevi, natrijevi in kloridni ioni prehajajo skozi membrano hkrati (s prepustnostjo P K, P Na in P Cl oziroma). Domneva se, da potencial enakomerno pada v membrani, tako da je poljska jakost konstantna. V tem primeru velja Goldmanova enačba ali enačba konstantnega polja :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Za večino celičnih membran P K približno 30-krat višji od R Na (glejte tudi razdelek 1.3). Relativna vrednost PCl močno se razlikuje; za številne membrane PCl majhna v primerjavi z R K , vendar za druge (npr. v skeletnih mišicah) PCl , veliko višje od R K.
Aktivni transport, natrijeva črpalka . Prejšnji razdelek opisuje pasivno difuzijo ionov in posledično membranski potencial pri danih koncentracijah znotraj in zunajceličnih ionov. Vendar se zaradi tega procesa koncentracija ionov v celici ne stabilizira samodejno, saj membrana
potencial je nekoliko bolj elektronegativen kot E K , in še veliko več v primerjavi z E Na (približno +60 mV). Zaradi difuzije bi se morale znotrajcelične koncentracije ionov, vsaj kalija in natrija, izenačiti z zunajceličnimi. Stabilnost ionskega gradienta se doseže z aktivnim transportom: membranski proteini prenašajo ione čez membrano proti električnim in (ali) koncentracijskim gradientom, pri čemer za to porabijo presnovno energijo. Najpomembnejši proces aktivnega transporta je delo Na/K - črpalka, ki obstaja v skoraj vseh celicah;
črpalka črpa natrijeve ione iz celice, hkrati pa v celico črpa kalijeve ione. To zagotavlja nizko znotrajcelično koncentracijo natrijevih ionov in visoko kalija (tabela 1.1). Gradient koncentracije natrijevih ionov na membrani ima posebne funkcije, povezane s prenosom informacij v obliki električnih impulzov (glej razdelek 2.2), pa tudi z vzdrževanjem drugih aktivnih transportnih mehanizmov in regulacije volumna celic (glej spodaj). Zato ni presenetljivo, da se več kot 1/3 energije, ki jo porabi celica, porabi za Na / K črpalko, v nekaterih najbolj aktivnih celicah pa se za njeno delovanje porabi tudi do 70 % energije.
Transportni protein Na/K je ATPaza. Na notranji površini membrane razgradi ATP na ADP in fosfat (slika 1.6). Energija ene molekule ATP se porabi za transport treh natrijevih ionov iz celice in hkrati dveh kalijevih ionov v celico, kar pomeni, da se skupaj v enem ciklu iz celice odstrani en pozitiven naboj. Tako je črpalka Na/K elektrogeni(ustvarja električni tok skozi membrano), kar vodi do povečanja elektronegativnosti membranskega potenciala za približno 10 mV. Transportni protein izvaja to operacijo z visoko hitrostjo: od 150 do 600 natrijevih ionov na sekundo. Zaporedje aminokislin transportnega proteina je znano, vendar mehanizem tega kompleksnega izmenjevalnega transporta še ni jasen. Ta proces je opisan z uporabo energijskih profilov prenosa natrijevih ali kalijevih ionov z beljakovinami (slika 1.5.5). Po naravi spremembe teh profilov, povezanih s stalnimi spremembami konformacije transportnega proteina (proces, ki zahteva energijo), lahko sodimo o stehiometriji izmenjave: dva kalijeva iona se zamenjata za tri natrijeve ione.
Na/K-črpalka, kot izolirana Na+ /K+-odvisna membranska ATPaza, ki jo specifično zavira srčni glikozid ouabain (strofantin). Ker je delovanje Na/K črpalke večstopenjska kemična reakcija, je tako kot vse kemične reakcije v veliki meri odvisna od temperature, kar

riž. 1.6.Shema Na/K-črpalke-ATPaze (potopljena v lipidni dvosloj plazemske membrane), ki v enem ciklu odvzame tri ione Na + iz celice proti potencialnim in koncentracijskim gradientom in v celico pripelje dva K iona + . Med tem procesom se ena molekula ATP razdeli na ADP in fosfat. Na diagramu je ATPaza prikazana kot dimer, sestavljen iz velike (funkcionalne) in majhne podenote; v membrani obstaja kot tetramer, ki ga tvorita dve veliki in dve majhni podenoti
prikazano na sl. 1.7. Tukaj je prikazan pretok natrijevih ionov iz mišičnih celic glede na čas; to je praktično enakovredno pretoku natrijevih ionov, ki ga posreduje delovanje Na/K črpalke, ker je pasivni tok natrijevih ionov proti koncentracijskim in potencialnim gradientom izjemno majhen. Če pripravek ohladimo za približno 18°C, se bo pretok natrijevih ionov iz celice hitro zmanjšal za 15-krat, takoj po segrevanju pa se bo vrnil na prvotno raven. Takšno zmanjšanje pretoka natrijevih ionov iz celice je nekajkrat večje od tistega, ki bi ustrezal temperaturni odvisnosti difuzijskega procesa ali preproste kemične reakcije. Podoben učinek opazimo, ko je presnovna energija izčrpana zaradi zastrupitve z dinitrofenolom (DNP) (slika 1.7.5). Zato pretok natrijevih ionov iz celice zagotavlja energijsko odvisna reakcija – aktivna črpalka. Druga značilnost črpalke, skupaj s pomembno temperaturno in energijsko odvisnostjo, je prisotnost stopnje nasičenosti (kot pri vseh drugih kemičnih reakcijah); to pomeni, da hitrost črpalke ne more naraščati v nedogled, ko se poveča koncentracija transportiranih ionov (slika 1.8). Nasprotno pa tok pasivno difuzijske snovi raste sorazmerno z razliko v koncentraciji v skladu z difuzijskim zakonom (enačbi 1 in 2).

riž. 1.7. A, B. Na aktivni promet + . Y-os: pretok radioaktivnega 24 Na + iz celice (imp./min). abscisna os:čas od začetka poskusa. AMPAK. Celico ohladimo z 18,3°C na 0,5°C; tok Na+ izstop iz celice v tem obdobju zavira. B. Zatiranje pretoka Na + iz celice z dinitrofenolom (DNF) v koncentraciji 0,2 mmol/l (s spremembami)
Poleg Na/K črpalke plazemska membrana vsebuje še vsaj eno črpalko - kalcij; ta črpalka črpa kalcijeve ione (Ca 2+) iz celice in sodeluje pri vzdrževanju njihove znotrajcelične koncentracije na izjemno nizki ravni (tabela 1.1). Kalcijeva črpalka je prisotna v zelo visoki gostoti v sarkoplazmatskem retikulumu mišičnih celic, ki kopičijo kalcijeve ione zaradi razgradnje molekul ATP (glej 4. poglavje).
Učinki Na/K črpalke na membranski potencial in prostornino celic . Na sl. 1.9 prikazuje različne komponente membranskega toka in prikazuje znotrajcelične koncentracije ionov, ki

riž. 1.8.Razmerje med hitrostjo transporta molekul in njihovo koncentracijo (na mestu vstopa v kanal ali na mestu vezave črpalke) med difuzijo skozi kanal ali med črpalnim transportom. Slednji nasiči pri visokih koncentracijah (maksimalna hitrost, Vmax ) vrednost na abscisi, ki ustreza polovici največje hitrosti črpalke ( Vmax /2), je ravnotežna koncentracija TO m

riž. 1.9.Diagram, ki prikazuje koncentracije Na+ , K+ in Cl- znotraj in zunaj celice ter načini, kako ti ioni prodirajo v celično membrano (preko specifičnih ionskih kanalov ali s pomočjo Na/K črpalke. Pri danih koncentracijskih gradientih se ravnotežni potenciali E Na, E K in E C l - enaki navedenim, membranski potencial Em = – 90 mV
zagotoviti njihov obstoj. Preko kalijevih kanalčkov opazimo tok kalijevih ionov navzven, saj je membranski potencial nekoliko bolj elektropozitiven od ravnotežnega potenciala za kalijeve ione. Skupna prevodnost natrijevih kanalčkov je veliko nižja od prevodnosti kalijevih kanalčkov; natrijevi kanali so odprti veliko manj pogosto kot kalijevi kanali v stanju mirovanja; vendar približno enako število natrijevih ionov vstopi v celico, kot jo zapustijo kalijevi ioni, ker so za difuzijo natrijevih ionov v celico potrebne velike koncentracije in potencialni gradienti. Na/K črpalka zagotavlja idealno kompenzacijo za pasivne difuzijske tokove, saj prenaša natrijeve ione iz celice in kalijeve ione vanj. Tako je črpalka elektrogena zaradi razlike v številu prenesenih nabojev v celico in iz nje, ki pri normalni hitrosti delovanja ustvari membranski potencial okoli 10 mV je bolj elektronegativen, kot če bi ga ustvarili samo pasivni ionski tokovi (glej enačbo 7). Posledično se membranski potencial približa potencialu kalijevega ravnotežja, kar zmanjša uhajanje kalijevih ionov. Na dejavnost/K-črpalka se uravnava z znotrajcelično koncentracijo natrijevih ionov. Hitrost črpalke se upočasni, ko se koncentracija natrijevih ionov, ki jih je treba odstraniti iz celice, zmanjša (slika 1.8), tako da delovanje črpalke in pretok natrijevih ionov v celico uravnovešata med seboj in ohranjata znotrajcelično koncentracijo. natrijevih ionov na ravni približno 10 mmol / l.
Za vzdrževanje ravnovesja med črpanjem in pasivnimi membranskimi tokovi je potrebnih veliko več molekul Na/K-črpalke kot je kanalskih proteinov za kalijeve in natrijeve ione. Ko je kanal odprt, skozenj v nekaj milisekundah preide na desetine tisoč ionov (glej zgoraj), in ker se kanal običajno odpre večkrat na sekundo, v tem času skozenj skupaj preide več kot 10 5 ionov. En protein črpalke premakne več sto natrijevih ionov na sekundo, zato mora plazemska membrana vsebovati približno 1000-krat več molekul črpalke kot kanalske molekule. Meritve kanalskih tokov v mirovanju so pokazale povprečno en kalijev in en natrijev odprt kanal na 1 µm 2 membrane; iz tega sledi, da mora biti v istem prostoru prisotnih približno 1000 Na/K molekul črpalke, t.j. razdalja med njima je v povprečju 34 nm; premer črpalnega proteina kot proteina kanala je 8–10 nm. Tako je membrana dovolj gosto nasičena s črpalnimi molekulami
Dejstvo, da se pretok natrijevih ionov v celico in kalijevih ionov iz celice kompenzira z delovanjem črpalke, ima še eno posledico, ki je v vzdrževanju stabilnega osmotskega tlaka in konstanten volumen. V notranjosti celice je visoka koncentracija velikih anionov, predvsem beljakovin (A - v tabeli 1.1), ki ne morejo prodreti skozi membrano (ali jo prodrejo zelo počasi) in so zato fiksna komponenta znotraj celice. Za uravnoteženje naboja teh anionov je potrebno enako število kationov. Zaradi delovanja Na/K črpalke so ti kationi predvsem kalijevi ioni. Do občutnega povečanja znotrajcelične koncentracije ionov bi lahko prišlo le ob povečanju koncentracije anionov zaradi pretoka C1 – vzdolž koncentracijskega gradienta v celico (tabela 1.1), vendar temu nasprotuje membranski potencial. Dohodni tok Cl- opazujemo le, dokler ni dosežen ravnotežni potencial za kloridne ione; to opazimo, ko je gradient kloridnih ionov skoraj nasproten gradientu kalijevih ionov, saj so kloridni ioni negativno nabiti (enačba 4). Tako se vzpostavi nizka znotrajcelična koncentracija kloridnih ionov, kar ustreza nizki zunajcelični koncentraciji kalijevih ionov. Rezultat je omejitev skupnega števila ionov v celici. Če membranski potencial pade, ko je črpalka Na/K blokirana, na primer med anoksijo, se ravnotežni potencial za kloridne ione zmanjša in temu primerno se poveča znotrajcelična koncentracija kloridnih ionov. Ob vzpostavitvi ravnovesja nabojev v celico vstopijo tudi kalijevi ioni; skupna koncentracija ionov v celici se poveča, kar poveča osmotski tlak; to prisili vodo, da vstopi v celico. Celica nabrekne. Ta oteklina se vidi in vivo v pogojih pomanjkanja energije.
koncentracijski gradient Na + kot gonilna sila za membranski transport . Pomen Na/K– črpalke za celico ni omejen na stabilizacijo normalnih gradientov K+ in Na+ čez membrano. Energija, shranjena v membranskem gradientu Na+ , se pogosto uporablja za zagotavljanje membranskega transporta za druge snovi. Na primer, na sl. 1.10 prikazuje "symport" Na+ in molekule sladkorja v celico. Membranski transportni protein prenaša molekulo sladkorja v celico tudi proti koncentracijskemu gradientu, medtem ko se Na + giblje vzdolž gradienta koncentracije in potenciala ter zagotavlja energijo za prevoz sladkorjev. Takšen transport sladkorjev je v celoti odvisen od obstoja visokega gradienta Na+ ; če je znotrajcelična koncentracija Na+ znatno poveča, nato se transport sladkorjev ustavi. Za različne c Akharov, obstajajo različni sistemi simbolov. Prevoz aminokislin v kletki, podobni transportu c akharov, prikazan na sl. 1,10; zagotavlja ga tudi gradient Na+ , obstaja vsaj pet različnih sistemov symport, od katerih je vsak specializiran za eno skupino sorodnih aminokislin.
Poleg sistemov symport obstajajo tudi "protiport". Eden od njih, na primer, v enem ciklu prenese en kalcijev ion iz celice v zameno za tri prihajajoče natrijeve ione (slika 1.10). Energija za transport Ca 2+ nastane zaradi vstopa treh natrijevih ionov vzdolž gradienta koncentracije in potenciala. Ta energija zadostuje (pri potencialu mirovanja) za vzdrževanje visokega gradienta kalcijevih ionov (od manj kot 10–7 mol/l znotraj celice do približno 2 mmol/l zunaj celice).
Endo- in eksocitoza . Za nekatere snovi, ki vstopijo v celico ali jih je treba odstraniti

riž. 1.10.Beljakovine, vgrajene v lipidni dvosloj membrane, posredujejo pri uvozu glukoze in Na+ v celico, kot tudi Ca2+/Na+ -antiport, pri katerem je gonilna sila gradient Na + na celični membrani
od nje ni transportnih kanalov; take snovi vključujejo na primer beljakovine in holesterol. Lahko prehajajo skozi plazemsko membrano v vezikli oz mehurčki, z endo- in eksocitozo. Na sl. 1.11 prikazuje glavne mehanizme teh procesov. Med eksocitozo določene organele (glej spodaj) tvorijo vezikle, napolnjene s snovjo, ki jo je treba odstraniti iz celice, kot so hormoni ali zunajcelični encimi. Ko takšni mehurčki dosežejo plazemsko membrano, se njihova lipidna membrana zlije z njo in tako omogoči, da vsebina uide v zunanje okolje. Pri nasprotnem procesu, endocitozi, plazemska membrana invaginira, tvori jamo, ki se nato poglobi in zapre ter tako nastane znotrajcelični mehurček, napolnjen z zunajcelično tekočino in nekaterimi makromolekulami. Da bi zagotovili to membransko fuzijo in zapiranje vezikla, delujejo kontraktilni elementi citoskeleta v povezavi s samimi membranami (glej spodaj). Endocitoza ne vključuje vedno preprostega zajema zunajceličnega medija v celico. Celična membrana vsebuje, pogosto organizirane v specializirane skupine, specifične receptorje za makromolekule, kot so insulin ali antigeni. Ko se te makromolekule vežejo na svoje receptorje, pride do endocitoze v predelu membrane, ki obdaja receptor, in makromolekula se selektivno transportira v celico (slika 1.12, B).
V celicah se nenehno pojavljata endo- in eksocitoza. Količina membranskega materiala, ki kroži, je pomembna; v 1 uri makrofag absorbira v obliki veziklov dvakratno površino svoje citoplazemske membrane. V večini celic promet membranskega materiala ni tako intenziven, vendar bi moral biti še vedno pomemben.

riž. 1.11.eksocitoza in endocitoza. gor: znotrajcelični mehurček se zlije z lipidnim dvoslojem plazemske membrane in se odpre v zunajcelični prostor. Ta proces se imenuje eksocitoza. na dnu: plazemska membrana invaginira na majhnem območju in zapre mehurček, napolnjen z zunajceličnim materialom. Ta proces se imenuje endocitoza.
1.3. Prenos snovi znotraj celice
Endo- in eksocitoza nista samo procesa transporta snovi skozi celično membrano, temveč tudi procesi membranske izmenjave - strukturnih sestavin same celice. Predmet obravnave v tem razdelku so drugi podobni transportni procesi v celici in njenih organelah.

riž. 1.12. A-B. Shema procesov, vključno z ekso- in endocitozo. AMPAK. Protein, sintetiziran v granularnem endoplazmatskem retikulumu, se preko Golgijevega aparata transportira do plazemske membrane, kjer se izloča z eksocitozo. B. Holesterol, vezan na delce LDL (lipoproteinov nizke gostote), se veže na plazemsko membrano, povzroči nastanek endocitnega mehurčka v tem predelu membrane in se transportira v lizosome, kjer se sprosti. IN. Zunajcelični material, zajet med endocitozo (na sliki na desni), se prenaša skozi celico v mehurčkih ali mehurčkih in se sprosti z eksocitozo (na sliki levo)
Difuzija . Seveda se v citosolu razlika v koncentraciji odpravi z difuzijo; enako velja za tekočine, ki jih vsebujejo organele. Zaradi visoke koncentracije raztopljenih beljakovin je difuzija tukaj veliko počasnejša kot v vodi. Lipidne membrane - okoli celice in znotraj organelov - so dvodimenzionalne tekočine, v katerih poteka difuzija. Lipidi v membranskem dvosloju se razpršijo znotraj lastne plasti, redko se premikajo iz enega v drugega. Beljakovine, ki so potopljene vanje, so tudi precej mobilne; vrtijo se okoli osi, pravokotne na membrano, ali difundirajo bočno z zelo različnimi difuzijskimi konstantami, 2–10.000-krat počasnejšimi kot fosfolipidi. Torej, če se nekateri proteini prosto gibljejo v lipidni plasti in z enako hitrostjo kot same lipidne molekule, potem so druge zasidrane, t.j. zelo močno povezana s citoskeletom. V membrani so "trajni" agregati specifičnih beljakovin, kot so pred- in postsinaptične strukture živčnih celic. Prosto gibljive beljakovine je mogoče dokazati tako, da jih vežemo na fluorescenčna barvila, ki se s kratkim osvetlitvijo majhnega območja membrane s kratkimi utripi inducirajo v žarenje. Takšni poskusi kažejo, da so beljakovine, vezane na barvilo, v manj kot 1 minuti enakomerno porazdeljene po membrani na razdaljah do 10 μm.
Aktivni transport v membranah organelov .
Procesi aktivnega transporta, ki igrajo ključno vlogo pri delovanju plazemske membrane, potekajo tudi znotraj celice, v membranah organelov. Specifične vsebine različnih organelov nastanejo deloma z notranjo sintezo in deloma z aktivnim transportom iz citosola. Eden od primerov slednjega je zgoraj omenjena črpalka Ca 2+ v sarkoplazmatskem retikulumu mišičnih celic. Posebej zanimivo je, da v primeru sinteze ATP v mitohondrijih velja nasprotno načelo, kar se dogaja v ATPaznih črpalkah plazemske membrane (slika 1.6). Med sintezo ATP oksidativni metabolizem vodi v nastanek strmega gradienta H+ na notranjih membranah. Ta gradient je gonilna sila za proces, ki je obraten od črpalnega cikla aktivnega molekularnega transporta: ioni H + se premikajo po membrani vzdolž gradienta, energija, ki se pri tem sprosti, pa zagotavlja sintezo ATP iz ADP in fosfata. . Nastali ATP pa celici zagotavlja energijo, tudi za aktivni transport.
Prevoz v mehurčkih . Celica ima veliko število organelov in pripadajočih veziklov (slika 1.1). Te organele, zlasti vezikli, so v stalnem gibanju in prenašajo svojo vsebino do drugih organele ali do plazemske membrane. Vezikli lahko migrirajo tudi iz celične membrane v organele, kot pri endocitozi.
proces izločanje beljakovin prikazano na sl. 1.12 AMPAK. Protein se sintetizira v bližini celičnega jedra na ribosomih, povezanih z endoplazmatskim retikulumom (tako imenovani zrnati ali grobi endoplazmatski retikulum); Ko pride v endoplazmatski retikulum, se protein pakira v transportne vezikle, ki se ločijo od organele in migrirajo v Golgijev aparat. Tu se združijo z rezervoarji Golgijevega aparata, kjer se protein modificira (tj. pretvori v glikoprotein). Na koncih cistern se mehurčki ponovno ločijo. Sekretorni vezikli, ki nosijo modificiran protein, se premikajo proti plazemski membrani in z eksocitozo sprostijo svojo vsebino.
Drug primer transportne poti v celici je prikazan na sl. 1,12, B; je privzem holesterola v celico. Holesterol, ki se prenaša v krvi, je v glavnem povezan z beljakovinami, kot so delci "lipoproteini nizke gostote"(LNP). Ti delci se pritrdijo na specifična mesta receptorja LDL na membrani, kjer se pojavi endocitoza in se LDL transportira v celico v "prevlečenih" mehurčkih. Ti mehurčki se zlijejo v endosome in med tem procesom izgubijo "zvonjenje". Endosomi se zlijejo s primarnimi lizosomi, ki vsebujejo pretežno hidrolitične encime, da tvorijo sekundarne, večje lizosome. V njih se holesterol sprosti iz delcev LDL in difundira v citosol, kjer postane dostopen na primer za sintezo lipidnih membran. Od endosomov se ločijo tudi vezikule, ki ne vsebujejo LDL, ki se na poseben način premikajo do plazemske membrane in se z njo zlijejo ter vračajo material membrane in verjetno LDL receptorje. Od trenutka, ko se delec LDL veže na membrano, mine 10–15 minut, dokler se holesterol ne sprosti iz sekundarnega lizosoma. Motnje pri vezavi in absorpciji LDL, torej pri oskrbi celice s holesterolom, imajo odločilno vlogo pri razvoju resne in razširjene bolezni, ateroskleroze (»otrdelosti« arterij).
Obstaja veliko drugih transportnih poti, podobnih tistim, prikazanim na sl. 1.11 in 1.12,A, s pomočjo katerih se v celici premikajo specifični vezikli. Ni natančno znano, kako se premikajo, a verjetno so v ta proces vključeni elementi citoskeleta. Vezikli lahko drsijo vzdolž mikrotubulov, v tem primeru se zdi, da energijo za gibanje zagotavlja beljakovina, povezana z vezikli, ATPaza (glej spodaj). Popolnoma nerazumljivo ostaja, koliko različnih veziklov, ki se premikajo drug za drugim v vse smeri, pride do cilja. Očitno jih je treba "označiti" tako, da jih prometni sistem prepozna in pretvori v namensko gibanje.
Transport s tvorbo in uničenjem organelov . Do sedaj smo endo- in eksocitozo obravnavali kot procese transporta vsebine veziklov. Obstaja še en vidik teh procesov, ki je sestavljen iz dejstva, da usmerjeno odstranjevanje plazemske membrane na enem področju celične površine z endocitozo in, nasprotno, njeno dodajanje drugemu z eksocitozo, premakne pomembne dele celične membrane. membrano (slika 1.12.E), ki daje celici možnost, da na primer tvori izrast ali se premakne.
Podobne preureditve so značilne tudi za citoskelet, zlasti za mikrofilamente in mikrotubule (slika 1.1). Mikrofilamenti sestavljen predvsem iz F-aktin protein ki se lahko sestavi v vlaknaste snope zaradi polimerizacije monomera iz citosola. Snopi so polarizirani, kar pomeni, da pogosto rastejo samo z enega konca in nabirajo nove aktinske molekule, drugi konec pa je inerten ali pa pride do razstavljanja. Zaradi te polarizirane rasti se mikrofilamenti učinkovito premikajo in struktura njihove mreže se lahko spremeni. Prehod aktina iz depolimeriziranega stanja (sol) v organizirano stanje (gel) se lahko zgodi zelo hitro pod vplivom drugih beljakovin ali sprememb v koncentraciji ionov (glej spodaj). Obstajajo tudi beljakovine, zaradi katerih se aktinski filamenti razgradijo na kratke fragmente. Tanki izrastki številnih celic – filopodij – vsebujejo osrednji snop aktina (slika 1.1), različni premiki filopodij pa so verjetno posledica aktinskih prehodov: polimerizacija – depolimerizacija.
mikrotubuletudi pogosto podvrženi podobnim gibom. Mehanizem teh premikov je podoben - polimerizacija tubulina iz citosola tako, da eden od koncev mikrotubule raste, drugi pa se bodisi ne spremeni ali pa se tam razstavi. Tako se lahko mikrotubul z ustreznim dodajanjem ali odstranitvijo materiala premika skozi citosol.
Aktivna gibanja citoskeleta . Spremembe v citoskeletnih strukturah se lahko pojavijo kot posledica tako aktivnih premikov kot preureditev, opisanih zgoraj. V mnogih primerih gibanje mikrotubulov in aktinskih filamentov poganjajo kontraktilni proteini, ki vežejo filamente ali tubule in jih lahko premikajo drug glede drugega. veverice miozin in dinein prisoten v citosolu vseh celic v relativno visokih koncentracijah; so elementi, ki pretvarjajo energijo v gibanje v specializiranih celicah (mišice) in organele (cilije). V mišičnih celicah miozin tvori debele filamente, usmerjene vzporedno z aktinskimi filamenti. Molekula miozina se s svojo "glavo" pritrdi na aktinski filament in z uporabo energije ATP premakne miozin vzdolž molekule aktina. Miozin se nato loči od aktina. Nabor številnih takih ciklov povezava-odklop vodi do makroskopskega krčenje mišičnih vlaken(4. poglavje). Dynein ima podobno vlogo pri gibanju mikrotubulov med delovanjem cilije (slika 1.1). V citoplazmi nespecializiranih celic miozin in dinein ne tvorita običajnih vlaken, temveč v večini primerov majhne skupine molekul. Tudi v obliki tako majhnih agregatov so sposobni premikati aktinske filamente ali mikrotubule. riž. 1.13 ponazarja ta proces, ko so nasprotno polarizirane molekule miozina pritrjene tudi na dva aktinska filamenta, polarizirana v nasprotnih smereh. Glavne skupine miozina se upognejo proti repu molekule, ki porabljajo ATP, dva aktinska filamenta pa se premakneta v nasprotni smeri, nakar se miozin loči od njiju. Tovrstna gibanja, med katerimi se energija ATP pretvori v mehansko delo, lahko spremenijo obliko citoskeleta in posledično celic ter zagotovijo transport organelov, povezanih s citoskeletom.
Na aksonu živčne celice lahko najbolj jasno pokažemo procese znotrajceličnega transporta. transport aksonov tukaj podrobno razpravljamo, da ponazorimo dogodke, ki se verjetno zgodijo na podoben način v večini celic. Akson s premerom le nekaj mikronov je lahko dolg en meter ali več in trajalo bi leta, da bi se proteini z difuzijo premaknili od jedra do distalnega konca aksona. Že dolgo je znano, da ko se kateri koli del aksona zoži, se proksimalni del aksona razširi. Videti je, da je centrifugalni tok blokiran v aksonu. Takšne tok – hiter transport aksonov lahko se dokaže s premikom radioaktivnih markerjev, kot v poskusu, prikazanem na sl. 1.14. Radioaktivno označen levcin smo injicirali v ganglij dorzalne korenine, nato pa smo od 2. do 10. ure merili radioaktivnost v ishiasnem živcu na razdalji 166 mm od teles nevronov. 10 ur se vrh radioaktivnosti na mestu injiciranja ni bistveno spremenil. Toda radioaktivni val se je širil vzdolž aksona s konstantno hitrostjo približno 34 mm na 2 uri ali 410 mm/dan. Dokazano je, da v vseh nevronih homoiotermnih živali hiter transport aksonov poteka z enako hitrostjo in ni opaznih razlik med tankimi, nemieliniziranimi vlakni in najdebelejšimi aksoni, pa tudi med motoričnimi in senzoričnimi vlakni. Vrsta radioaktivnega markerja prav tako ne vpliva na hitrost hitrega aksonskega transporta; Kot markerji lahko služijo različne radioaktivne snovi.

riž. 1.13.Nemišični miozinski kompleks z določeno orientacijo se lahko veže na aktinske filamente različne polarnosti in jih z uporabo energije ATP premakne drug glede drugega.
molekule, kot so različne aminokisline, ki so vključene v telesne beljakovine nevrona. Če analiziramo periferni del živca, da ugotovimo naravo prenašalcev radioaktivnosti, ki se tu prenaša, potem takšne nosilce najdemo predvsem v beljakovinski frakciji, pa tudi v sestavi mediatorjev in prostih aminokislin. Ker vemo, da so lastnosti teh snovi različne in predvsem velikosti njihovih molekul različne, lahko konstantno hitrost transporta razložimo le s transportnim mehanizmom, ki je vsem skupen.
Opisano zgoraj hiter transport aksonov je anterogradni tj. usmerjeno stran od celičnega telesa. Dokazano je, da se nekatere snovi premikajo z obrobja v celično telo s pomočjo retrogradni transport. Na primer, acetilholinesteraza se transportira v tej smeri s hitrostjo, ki je dvakrat nižja od hitrosti hitrega aksonskega transporta. Retrogradno se premika tudi marker, ki se pogosto uporablja v nevroanatomiji - peroksidaza iz hrena. Retrogradni transport ima verjetno pomembno vlogo pri uravnavanju sinteze beljakovin v celičnem telesu. Nekaj dni po transekciji aksona v telesu celice opazimo kromatolizo, kar kaže na kršitev sinteze beljakovin. Čas, potreben za kromatolizo, je povezan s trajanjem retrogradnega transporta od mesta transekcije aksona do telesa celice. Takšen rezultat nakazuje tudi razlago za to kršitev - moten je prenos z obrobja "signalne snovi", ki uravnava sintezo beljakovin. Očitno so glavna "vozila", ki se uporabljajo za hitro aksonalno
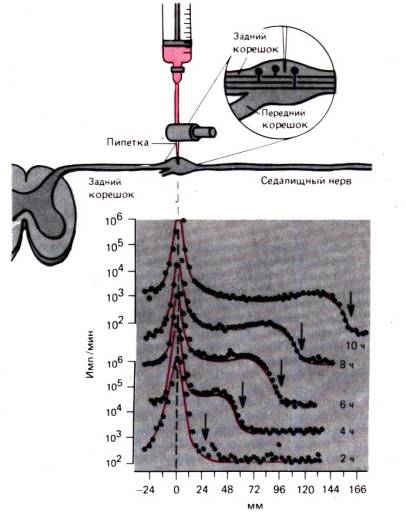
riž. 1.14.Poskus, ki dokazuje hiter transport aksonov v senzoričnih vlaknih ishiadičnega živca mačke. Levcin, označen s tritijem, se injicira v ganglij hrbtne korenine, radioaktivnost v gangliju in senzoričnih vlaknih pa se izmeri 2, 4, 6, 8 in 10 ur po injiciranju. (spodaj slike). Avtor abscisa razdalja od ganglija do odsekov ishiadičnega živca, kjer se meritev, se odloži. Na osi y je samo za zgornjo in spodnjo krivuljo v logaritmični lestvici izrisana radioaktivnost (imp./min). "Val" povečane radioaktivnosti (puščice) premika se s hitrostjo 410 mm/dan (za )
prevoz so vezikli (vezikli) in organeli, kot so mitohondriji, ki vsebujejo snovi, ki jih je treba prevažati. Gibanje največjih veziklov ali mitohondrijev lahko opazujemo z mikroskopom in vivo . Takšni delci naredijo kratke, hitre premike v eno od smeri, se ustavijo, se pogosto premaknejo malo nazaj ali vstran, se ponovno ustavijo in nato naredijo sunk v glavni smeri. 410 mm/dan ustreza povprečni anterogradni hitrosti približno 5 μm/s; hitrost vsakega posameznega gibanja bi morala biti zato veliko večja, in če upoštevamo velikost organelov, filamentov in mikrotubul, potem so ti premiki res zelo hitri. Hiter transport aksonov zahteva znatno koncentracijo ATP. Strupi, kot je kolhicin, ki uničuje mikrotubule, prav tako blokirajo hiter aksonski transport. Iz tega sledi, da se v transportnem procesu, ki ga obravnavamo, mehurčki in organele premikajo po mikrotubulih in aktinskih filamentih; to gibanje zagotavljajo majhni agregati molekul dineina in miozina, ki delujejo, kot je prikazano na sl. 1.13, z uporabo energije ATP.
Vključen je lahko tudi hiter transport aksonov patološki procesi. Nekateri nevrotropni virusi (na primer virusi herpesa ali otroške paralize) prodrejo v akson na periferiji in se s pomočjo retrogradnega transporta premaknejo v nevronsko telo, kjer se razmnožujejo in izvajajo svoj toksični učinek. Tetanusni toksin, beljakovino, ki jo proizvajajo bakterije, ki vstopijo v telo s kožnimi lezijami, prevzamejo živčni končiči in se prenesejo v telo nevronov, kjer povzroči značilne mišične krče. Znani so primeri toksičnih učinkov na sam transport aksonov, na primer izpostavljenost industrijskemu topilu akrilamidu. Poleg tega se domneva, da patogeneza beriberi beriberi in alkoholne polinevropatije vključuje kršitev hitrega aksonskega transporta.
Poleg hitrega transporta aksonov v celici je tudi precej intenziven počasen transport aksonov. Tubulin se premika vzdolž aksona s hitrostjo približno 1 mm/dan, medtem ko se aktin premika hitreje, do 5 mm/dan. S temi komponentami citoskeleta migrirajo tudi drugi proteini; na primer, zdi se, da so encimi povezani z aktinom ali tubulinom. Hitrosti gibanja tubulina in aktina so približno skladne s stopnjami rasti, ugotovljenimi za prej opisani mehanizem, ko so molekule vključene v aktivni konec mikrotubule ali mikrofilamenta. Zato je ta mehanizem lahko osnova počasnega aksonskega transporta. Hitrost počasnega transporta aksonov približno ustreza tudi hitrosti rasti aksonov, kar očitno kaže na omejitve, ki jih struktura citoskeleta nalaga drugemu procesu.
Ob zaključku tega razdelka je treba poudariti, da celice nikakor niso statične strukture, kot se pojavljajo na primer na elektronskih mikroskopskih fotografijah. plazemska membrana in še posebej organele so v nenehnem hitrem gibanju in nenehnem prestrukturiranju; to je edini razlog, da lahko delujejo. Poleg tega to niso preproste komore, v katerih potekajo kemične reakcije, ampak visoko organizirani konglomerati membran in vlaken, pri katerem reakcije potekajo v optimalno organiziranem zaporedju.
1.4. Regulacija celičnih funkcij
Vzdrževanje posamezne celice kot funkcionalne enote v veliki meri ureja jedro; preučevanje takšnih regulativnih mehanizmov je predmet celične biologije in biokemije. Hkrati morajo celice spreminjati svoje funkcije v skladu z okoljskimi razmerami in potrebami drugih telesnih celic, torej služijo kot objekti funkcionalne regulacije. V nadaljevanju na kratko razmislimo, kako ti regulativni vplivi delujejo na plazemsko membrano in kako dosežejo znotrajcelične organele.
Regulativni učinki na celično membrano
Membranski potencial . V mnogih primerih se regulacija celičnih funkcij izvaja s spreminjanjem membranskega potenciala. Spremembe lokalnega potenciala so možne, ko: 1) skozi membrano teče tok iz sosednjega celičnega območja ali ga ustvari druga celica; 2) zunajcelična koncentracija ionov se spremeni (pogosto [K + ] ven ); 3) membranski ionski kanali se odprejo. Spremembe membranskega potenciala lahko vplivajo na konformacijo membranskih proteinov, kar povzroči odpiranje ali zapiranje kanalov. Kot je opisano zgoraj, je delovanje nekaterih membranskih črpalk odvisno od membranskega potenciala. Živčne celice so specializirane za zaznavanje sprememb membranskega potenciala kot informacije, ki jih je treba obdelati in prenesti (glej 2. poglavje).
zunajcelične regulatorne snovi . Najpomembnejši regulacijski mehanizem, ki vključuje zunajcelične snovi, je njihova interakcija s specifičnimi receptorji na plazemski membrani ali znotraj celice. Te snovi vključujejo sinaptične mediatorje, ki prenašajo informacije med živčnimi celicami, lokalnimi sredstvi in snovmi, ki krožijo v krvi in dosežejo vse celice telesa, kot so hormoni in antigeni. sinaptični nevrotransmiterji so majhne molekule, sproščene iz živčnih končičev na sinapsi;
ko dosežejo plazemsko membrano sosednje postsinaptične celice, sprožijo električne signale ali druge regulacijske mehanizme. To vprašanje je podrobno obravnavano v poglavju. 3.
Lokalna kemična sredstva pogosto izločajo specializirane celice. Prosto difundirajo v zunajceličnem prostoru, vendar je njihovo delovanje omejeno na majhno skupino celic zaradi hitrega uničenja teh snovi, bodisi spontano bodisi z delovanjem encimov. En primer sproščanja takih sredstev je sproščanje histamin mastociti ob poškodbi ali imunskem odzivu. Histamin povzroča sprostitev gladkih mišičnih celic žil, povečuje prepustnost žilnega endotelija in stimulira senzorične živčne končiče, ki posredujejo občutek srbenja. Druge lokalne kemične snovi izločajo številne druge celice. Tipični lokalni agenti so prostaglandini, ki sestavljajo skupino približno 20 derivatov maščobnih kislin. Kontinuirano se sproščajo iz široko razširjenih celic, vendar delujejo le lokalno, saj jih membranske fosfolipaze hitro uničijo. Različni prostaglandini imajo širok spekter delovanja: lahko sprožijo krčenje gladkih mišičnih celic, povzročijo agregacijo krvnih ploščic (trombocitov) ali zavirajo razvoj rumenega telesa v jajčnikih.
Drugi lokalni agenti služijo rastni dejavniki. Najbolj znan živčni rastni faktor (NGF) za simpatične nevrone, ki je nujen za rast in preživetje teh nevronov med razvojem in vivo ali v celični kulturi. Očitno ciljne celice za ta razred nevronov izločajo NGF in tako zagotavljajo pravilno inervacijo. Pri oblikovanju organov morajo celice pogosto "najti pot" do ciljnih celic, ki se lahko nahajajo na precejšnji razdalji. V skladu s tem mora obstajati veliko specializiranih rastnih faktorjev, kot je NGF.
Hormoni in antigeni prenaša kri do vseh celic. Antigeni izzovejo imunski odziv celic, ki nosijo specifična protitelesa. Antigeni pa so praviloma tuje snovi, ki ne nastajajo v reagirajočem organizmu (za več podrobnosti glej 18. poglavje). Nekateri hormoni, kot sta inzulin ali tiroksin, vplivajo na najrazličnejše vrste celic, medtem ko drugi, kot so spolni hormoni, vplivajo le na določene vrste celic. Hormoni so bodisi peptidi, katerih delovanje sproži njihova vezava na receptor na celični membrani, bodisi steroidi in tiroksin, ki difundirajo skozi lipidno membrano in se vežejo na znotrajcelične receptorje. Steroidni hormoni se vežejo na jedrski kromatin, kar povzroči prepisovanje določenih genov. Posledično proizvedene beljakovine povzročajo spremembo celičnih funkcij, kar je specifičen učinek hormonov. Vprašanja, povezana s sproščanjem in delovanjem hormonov, so podrobno obravnavana v pogl. 17.
Znotrajcelična komunikacija, ki vključuje sekundarne selce
Zgoraj opisane regulacijske funkcije vključujejo učinke na celično membrano. Informacije, ki jih prejme celična membrana, morajo pogosto povzročiti reakcijo v organelah in jih do njih prenašajo različne snovi, znane kot drugi glasniki (v nasprotju s prvimi, ki prihajajo v celico iz zunanjih virov). Študija drugih mediatorjev se hitro razvija in ni zagotovila, da bo trenutna raven razumevanja problema dovolj popolna. Tu se bomo dotaknili treh dobro raziskanih mediatorjev: Ca 2+, cAMP in inozitol trifosfata.
kalcij.Najpreprostejši znotrajcelični mediator je ion Ca 2+. Njegova prosta koncentracija v celici v mirovanju je zelo nizka in znaša 10 -8 -10 -7 mol/l. V celico lahko vstopi skozi specifične membranske kanale, ko so odprti, na primer, ko se spremeni membranski potencial (glej 2. poglavje). Posledično povečanje Ca 2+ sproži pomembne reakcije v celici, kot je krčenje miofibril, ki je osnova za krčenje mišic (glej 4. poglavje), ali sproščanje veziklov, ki vsebujejo nevrotransmiterje, iz živčnih končičev (glej poglavje 3). . Obe reakciji zahtevata koncentracijo Ca 2+ približno 10–5 mol/l. Ca 2+, ki ima regulacijski učinek, se lahko sprosti tudi iz znotrajceličnih depojev, kot je endoplazmatski retikulum. Sprostitev Ca 2+ iz depoja zahteva sodelovanje drugih posrednikov (glej npr. sliko 1.16).
Ciklični adenozin monofosfat, cAMP. V zadnjem času je bilo dokazano, da je ciklični adenozin monofosfat (cAMP), derivat glavnega vira energije v telesu, ATP, pomemben drugi glasnik. Kompleksna veriga reakcij, prikazana na sl. 1.15, se začne od receptorja Rs na zunanji površini plazemske membrane, ki lahko služi kot specifično vezavno mesto za različne mediatorje in hormone. Po vezavi na specifično "stimulativno" molekulo Rs spremeni svojo konformacijo; te spremembe vplivajo na beljakovine Gs na notranji površini membrane tako, da je mogoče slednjo aktivirati z intracelularnim gvanozin trifosfatom (GTP). aktivirane beljakovine Gs , po drugi strani stimulira encim na notranji površini membrane, adenilat ciklazo (AC), ki katalizira tvorbo cAMP iz ATP. V vodi topen cAMP in je posrednik, ki prenaša učinek

riž. 1.15.Veriga reakcij, ki vključuje znotrajcelični mediator cAMP (ciklični adenozin monofosfat). Ekscitatorni ali zaviralni zunanji signali aktivirajo membranske receptorje R s ali Ri . Ti receptorji uravnavajo proces vezave G -proteini z znotrajceličnim GTP (gvanozin trifosfat), s čimer stimulirajo ali zavirajo intracelularno adenilat ciklazo (AC). Encim za ojačanje AC pretvori adenozin trifosfat (ATP) v cAMP, ki se nato s pomočjo fosfodiestra razcepi na AMP. Prosti cAMP difundira v celico in aktivira adenilat kinazo (A-kinazo), pri čemer se sprosti njena katalitska podenota C, ki katalizira fosforilacijo znotrajceličnih beljakovin, t.j. tvori končni učinek zunajceličnega dražljaja. Shema prikazuje tudi farmakološka zdravila in toksine, ki sprožijo (+) ali zavirajo (-) nekatere reakcije (vendar s spremembami)
stimulacija zunajceličnega receptorja Rs na notranje strukture celice.
Vzporedno s stimulativno verigo reakcij, ki vključujejo Rs možna vezava inhibitornih mediatorjev in hormonov na ustrezen receptor R i kar spet preko GTP-aktiviranega proteina G , zavira proizvodnjo AC in s tem cAMP. Z razpršitvijo v celico cAMP reagira z adenilat kinazo (A-kinazo); S tem se sprosti podenota C, ki katalizira fosforilacijo beljakovin. Ta fosforilacija pretvori beljakovine v njihovo aktivno obliko in zdaj lahko izvajajo svoje specifično regulacijsko delovanje (na primer povzročijo razgradnjo glikogena). Ta kompleksen regulacijski sistem je izjemno učinkovit, saj je končni rezultat fosforilacija številnih beljakovin, torej regulatorni signal prehaja skozi verigo z velikim faktorjem ojačanja. Zunanji mediatorji, ki se vežejo na receptorje R s in R i specifične za vsako od njih so izjemno raznolike. Adrenalin, zafrkavanje R s ali R i sodeluje pri uravnavanju presnove lipidov in glikogena, pa tudi pri povečanem krčenju srčne mišice in pri drugih reakcijah (glej 19. poglavje). ščitnični stimulirajoči hormon, aktiviranje Rs , spodbuja izločanje hormona tiroksina s strani ščitnice, prostaglandin I pa zavira agregacijo trombocitov. Zaviralni učinki, vključno z adrenalinom, posredovani R i izraženo v upočasnitvi lipolize. V to smer, sistem cAMP je večnamenski znotrajcelični regulacijski sistem, ki jih je mogoče natančno nadzorovati z zunajceličnimi stimulacijskimi in zaviralnimi signalnimi sredstvi.
Inozitol fosfat "IF h ". Intracelični sistem drugega mediatorja, inozitol fosfata, je bil odkrit šele pred kratkim (slika 1.16). V tem primeru ni zaviralne poti, obstaja pa podobnost s sistemom cAMP, pri katerem se učinek stimulacije R receptorja prenese na GTP-aktiviran G-protein na notranji površini membrane. Na naslednji stopnji se običajni membranski lipidni fosfatidilinozitol (PI), ki je prej prejel dve dodatni fosfatni skupini, pretvori v PI-difosfat (FIF 2), ki ga aktivirana fosfodiesteraza (PDE) razcepi v inozitol trifosfat(IFz) in lipid diacilglicerol(DAG). Inozitol trifosfat je vodotopen sekundarni prenašalec, ki difundira v citosol. Deluje predvsem tako, da sprošča Ca 2+ iz endoplazmatskega retikuluma. Ca 2+ pa deluje kot posrednik, kot je opisano zgoraj; na primer aktivira Ca2+-odvisno fosfokinazo, ki fosforilira encime. Lipidna podenota DAG (slika 1.16) prav tako prenaša signal, ki se v lipidni fazi plazemske membrane razprši do C-kinaze, ki se nahaja na njeni notranji površini, ki se aktivira s sodelovanjem fosfatidilserina kot kofaktorja. Nato C-kinaza sproži fosforilacijo beljakovin in jih pretvori v aktivno obliko.
Znotrajcelični sistem drugega vmesnega interferona 3 lahko nadzorujejo tudi različni zunanji mediatorji in hormoni, vključno z acetilholinom, serotoninom, vazopresinom in hormonom, ki stimulira ščitnico; tako kot sistem cAMP, so zanj značilni različni znotrajcelični učinki. Možno je, da se ta sistem aktivira tudi s svetlobo v vidnem receptorju očesa in ima osrednjo vlogo pri fototransdukciji (glej 11. poglavje). Prvič v individualnem razvoju organizma se receptor sistema IGF aktivira s semenčico, zaradi česar IGF sodeluje v regulacijskih reakcijah, ki spremljajo oploditev jajčeca.
Sistema cAMP in IFz-DAG sta zelo učinkovita biološki ojačevalci. so

riž. 1.16.Veriga reakcij, ki vključujejo znotrajcelični vmesni interferon (inozitol trifosfat). Tako kot v sistemu cAMP je zunajcelični signal posredovan prek proteinaG, ki v tem primeru aktivira fosfodiesterazo (PDE). Ta encim razgrajuje fosfatidilinozin difosfat (FIF) 2 ) v plazemski membrani pred IF h in diacilglicerol (DAG); ČE h difundira v citoplazmo. Tu povzroči sproščanje Ca 2+ iz endoplazmatskega retikuluma; povečanje koncentracije Ca 2+ v citoplazmi ([Ca 2+ ] i ) aktivira protein kinazo, ki fosforilira in zato aktivira encime. Drugi produkt, DAG, ostane v membrani in aktivira protein kinazo C (kofaktor-fosfatidilserin, PS). Protein kinaza C tudi fosforilira encime, ki posredujejo specifična dejanja, povezana s stimulacijo zunanjega receptorja. R . Veje verige reakcij, ki vključujejo IF h in DAG se lahko neodvisno aktivirata z ionomicinom oziroma forbolnim estrom (spremenjeno)
pretvori reakcijo med mediatorjem in receptorjem zunanje membrane v fosforilacijo številnih znotrajceličnih beljakovin, ki lahko nato vplivajo na različne celične funkcije. Eden od bistvenih vidikov problema je, da, kolikor danes vemo, obstajata le ta dva tesno povezana regulacijska sistema te vrste, ki ju uporabljajo številni zunanji mediatorji za uravnavanje različnih znotrajceličnih procesov. Hkrati ti regulativni sistemi, vključno s Ca 2+, tesno medsebojno delujejo, kar jim omogoča fino regulacijo celičnih funkcij.
Vadnice in vodniki
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts TO., Watson J.D.Molekularna biologija celice, New York in London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(ur.). biologija. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille IN. Ionski kanali ekscitabilnih membran. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(ur.). Biophysik. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermanna TO., Mahler H.Biokemija. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E.R., Schwartz-J.H.,(ur.). Principi nevronske znanosti, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., SchmidtW.Anatomski des Menschen. Berlin, Heidelberg, New York, Tokio, Springer, 1983.
Izvirni članki in ocene
8. Berridge M.J. Molekularna osnova komunikacije znotraj celice, sci. Amer 253 124 134 (1985).
9. Berridge M. J., Irvine R. F. Inozitol trifosfat, nov drugi posrednik pri prenosu celičnega signala. Narava, 312, 315 321 (1984).
10. Bretscher M.S. Molekule celične membrane, Sci. Amer., 253, 124–134 (1985).
11. Daut J.Živa celica kot stroj za pretvorbo energije. Minimalni model miokardnega metabolizma, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz IN. Vpliv natrijevih ionov na električno aktivnost velikanskega aksona lignjev. J Physiol. (Lon.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Aktivni transport kationov v velikanskih aksonih iz sepija in Loligo, J Physiol. (Lon.), 128, 28–42 (1955).
14. Daljša P. Ionski kanali s konformacijskimi podstanji, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Eksoplazmatski transport v normalnih in patoloških sistemih. V: Fiziologija in patologija aksonov, S.G. Waxman, Ed. New York, Raven Press, 1978.
Vabimo vas, da se seznanite z materiali in.
: celulozna membrana, membrana, citoplazma z organeli, jedro, vakuole s celičnim sokom.Prisotnost plastidov je glavna značilnost rastlinske celice.
Funkcije celične stene- določa obliko celice, ščiti pred okoljskimi dejavniki.
plazemska membrana- tanek film, sestavljen iz medsebojno delujočih lipidnih in beljakovinskih molekul, ločuje notranjo vsebino od zunanjega okolja, zagotavlja transport vode, mineralnih in organskih snovi v celico z osmozo in aktivnim prenosom ter odstranjuje tudi odpadne produkte.
citoplazma- notranje poltekoče okolje celice, v katerem se nahajajo jedro in organeli, zagotavlja povezave med njimi, sodeluje v glavnih procesih življenja.
Endoplazemski retikulum- mreža razvejanih kanalov v citoplazmi. Sodeluje pri sintezi beljakovin, lipidov in ogljikovih hidratov, pri transportu snovi. Ribosomi - telesa, ki se nahajajo na EPS ali v citoplazmi, sestavljena iz RNA in beljakovin, sodelujejo pri sintezi beljakovin. EPS in ribosomi so en sam aparat za sintezo in transport beljakovin.
mitohondrije-organele, ločene od citoplazme z dvema membranama. V njih se oksidirajo organske snovi in s sodelovanjem encimov se sintetizirajo molekule ATP. Povečanje površine notranje membrane, na kateri se nahajajo encimi zaradi krist. ATP je energijsko bogata organska snov.
plastidi(kloroplasti, levkoplasti, kromoplasti), je njihova vsebnost v celici glavna značilnost rastlinskega organizma. Kloroplasti so plastidi, ki vsebujejo zeleni pigment klorofil, ki absorbira svetlobno energijo in jo uporablja za sintezo organskih snovi iz ogljikovega dioksida in vode. Ločitev kloroplastov od citoplazme z dvema membranama, številnimi izrastki - grana na notranji membrani, v kateri se nahajajo molekule klorofila in encimi.
Golgijev kompleks- sistem votlin, ločenih od citoplazme z membrano. Kopičenje beljakovin, maščob in ogljikovih hidratov v njih. Izvajanje sinteze maščob in ogljikovih hidratov na membranah.
lizosomi- telesa, ločena od citoplazme z eno samo membrano. Encimi, ki jih vsebujejo, pospešujejo reakcijo cepitve kompleksnih molekul na preproste: beljakovine na aminokisline, zapletene ogljikove hidrate na preproste, lipide na glicerol in maščobne kisline ter uničujejo tudi odmrle dele celice, cele celice.
Vakuole- votline v citoplazmi, napolnjene s celičnim sokom, mesto kopičenja rezervnih hranil, škodljivih snovi; uravnavajo vsebnost vode v celici.
Jedro- glavni del celice, na zunanji strani prekrit z dvomembransko, prebodeno s porami jedrno ovojnico. Snovi vstopijo v jedro in se iz njega odstranijo skozi pore. Kromosomi so nosilci dednih informacij o značilnostih organizma, glavnih strukturah jedra, od katerih je vsaka sestavljena iz ene molekule DNK v kombinaciji z beljakovinami. Jedro je mesto sinteze DNK, i-RNA, r-RNA.

Prisotnost zunanje membrane, citoplazme z organeli, jedra s kromosomi.
Zunanja ali plazemska membrana- ločuje vsebino celice od okolja (druge celice, medcelična snov), sestoji iz lipidnih in beljakovinskih molekul, zagotavlja komunikacijo med celicami, transport snovi v celico (pinocitoza, fagocitoza) in iz celice.
citoplazma- notranje poltekoče okolje celice, ki zagotavlja komunikacijo med jedrom in organeli, ki se nahajajo v njej. Glavni procesi vitalne aktivnosti potekajo v citoplazmi.
celični organeli:
1) endoplazmatski retikulum (ER)- sistem razvejanih tubulov, ki sodelujejo pri sintezi beljakovin, lipidov in ogljikovih hidratov, pri transportu snovi v celici;
2) ribosomi- telesa, ki vsebujejo rRNA, se nahajajo na ER in v citoplazmi ter sodelujejo pri sintezi beljakovin. EPS in ribosomi so en sam aparat za sintezo in transport beljakovin;
3) mitohondrije- "elektrarne" celice, ločene od citoplazme z dvema membranama. Notranji tvori kriste (gube), ki povečajo njegovo površino. Encimi na kristah pospešujejo reakcije oksidacije organskih snovi in sintezo energijsko bogatih molekul ATP;
4) golgijev kompleks- skupina votlin, omejenih z membrano od citoplazme, napolnjenih z beljakovinami, maščobami in ogljikovimi hidrati, ki se bodisi uporabljajo v življenjskih procesih bodisi odstranijo iz celice. Membrane kompleksa izvajajo sintezo maščob in ogljikovih hidratov;
5) lizosomi- z encimi napolnjena telesa pospešujejo reakcije cepitve beljakovin na aminokisline, lipidov na glicerol in maščobne kisline, polisaharidov na monosaharide. V lizosomih se uničijo odmrli deli celice, cele celice in celice.
Vključki celic- Akumulacije rezervnih hranil: beljakovin, maščob in ogljikovih hidratov.
Jedro- najpomembnejši del celice. Pokrit je z dvojno membrano s porami, skozi katere nekatere snovi prodrejo v jedro, druge pa v citoplazmo. Kromosomi so glavne strukture jedra, nosilci dednih informacij o značilnostih organizma. Prenaša se v procesu delitve matične celice na hčerinske celice, z zarodnimi celicami pa na hčerinske organizme. Jedro je mesto sinteze DNK, mRNA, rRNA.
Naloga:
Pojasnite, zakaj se organele imenujejo specializirane strukture celice?
odgovor: organele imenujemo specializirane celične strukture, saj opravljajo strogo določene funkcije, dedne informacije so shranjene v jedru, ATP se sintetizira v mitohondrijih, fotosinteza poteka v kloroplastih itd.
Če imate vprašanja o citologiji, lahko zaprosite za pomoč pri
Tretja faza evolucije je videz celice.
Molekule beljakovin in nukleinskih kislin (DNK in RNA) tvorijo biološko celico, najmanjšo enoto življenja. Biološke celice so »gradniki« vseh živih organizmov in vsebujejo vse materialne kode razvoja.
Znanstveniki so dolgo časa menili, da je struktura celice izjemno preprosta. Sovjetski enciklopedični slovar razlaga koncept celice na naslednji način: "Celica je osnovni živi sistem, osnova zgradbe in življenja vseh živali in rastlin." Opozoriti je treba, da izraz "elementaren" nikakor ne pomeni "najpreprostejšega". Nasprotno, celica je edinstvena fraktalna stvaritev Boga, ki preseneča s svojo kompleksnostjo in hkrati z izjemno skladnostjo delo vseh njegovih elementov.
Ko smo s pomočjo elektronskega mikroskopa uspeli pogledati v notranjost, se je izkazalo, da je naprava najpreprostejše celice tako zapletena in nerazumljiva kot Vesolje samo. Danes je že ugotovljeno, da je "Celica posebna zadeva Vesolja, posebna zadeva Kozmosa." Ena sama celica vsebuje informacije, ki jih je mogoče vnesti v le nekaj deset tisoč zvezkov Velike sovjetske enciklopedije. tiste. celica je med drugim ogromen »biorezervoar« informacij.
Avtor sodobne teorije molekularne evolucije Manfred Eigen piše: »Da bi narava nastala po naključju, bi morala narava narediti približno 10130 poskusov in za to porabiti toliko molekul, ki bi zadostovalo za 1027 Vesolja.Če je bil protein zgrajen inteligentno, to je, da je bilo mogoče veljavnost vsake poteze preveriti s kakšnim izbirnim mehanizmom, je bilo potrebnih le okoli 2000 poskusov. Pridemo do paradoksnega zaključka: program za izgradnjo "primitivne žive celice" je kodiran nekje na ravni elementarnih delcev".
In kako bi bilo drugače. Vsaka celica, ki ima DNK, je obdarjena z zavestjo, se zaveda sebe in drugih celic ter je v stiku z vesoljem, saj je pravzaprav njegov del. In čeprav je število in raznolikost celic v človeškem telesu neverjetna (približno 70 bilijonov), so vse same sebi podobne, tako kot so vsi procesi, ki se dogajajo v celicah, sami sebi podobni. Po besedah nemškega znanstvenika Rolanda Glaserja je zasnova bioloških celic »zelo dobro premišljena«. Kdo je dobro premišljen?
Odgovor je preprost: beljakovine, nukleinske kisline, žive celice in vsi biološki sistemi so produkt ustvarjalne dejavnosti intelektualnega Stvarnika.
Zanimivo je: na atomski ravni ni razlik med kemično sestavo organskega in anorganskega sveta. Z drugimi besedami, na ravni atoma je celica ustvarjena iz istih elementov kot neživa narava. Razlike najdemo na molekularni ravni. V živih telesih so poleg anorganskih snovi in vode tudi beljakovine, ogljikovi hidrati, maščobe, nukleinske kisline, encim ATP sintaza in druge nizkomolekularne organske spojine.
Do danes je bila celica za namene študija dobesedno razstavljena na atome. Ni pa mogoče ustvariti vsaj ene žive celice, ker ustvariti celico pomeni ustvariti delček živega Vesolja. Akademik V.P. Kaznacheev meni, da je "celica kozmoplanetarni organizem ... Človeške celice so določeni sistemi eterično-torzijnih biotrkalnikov. V teh biotrkalnikih se dogajajo nam neznani procesi, materializacija kozmičnih oblik tokov, njihova kozmična transformacija in zaradi tega delci se materializirajo".
Voda.
Skoraj 80 % celične mase je voda. Po mnenju doktorja biologije S. Zenina je voda zaradi svoje gručaste strukture informacijska matrika za upravljanje biokemičnih procesov. Poleg tega je voda primarna "tarča", s katero delujejo nihanja zvočne frekvence. Urejenost celične vode je tako visoka (blizu urejenosti kristala), da jo imenujemo tekoči kristal.
veverice.
Beljakovine igrajo pomembno vlogo v biološkem življenju. Celica vsebuje več tisoč beljakovin, ki so edinstvene za to vrsto celice (z izjemo matičnih celic). Sposobnost sinteze lastnih beljakovin se podeduje od celice do celice in se ohranja vse življenje. V času življenja celice beljakovine postopoma spreminjajo svojo strukturo, njihova funkcija je oslabljena. Te izrabljene beljakovine se odstranijo iz celice in nadomestijo z novimi, zaradi česar se ohrani vitalna aktivnost celice.
Najprej opazimo gradbeno funkcijo beljakovin, saj so gradbeni material, ki sestavlja membrane celic in celičnih organelov, stene krvnih žil, kit, hrustanca itd.
Izredno zanimiva je signalna funkcija beljakovin. Izkazalo se je, da lahko beljakovine služijo kot signalne snovi, ki prenašajo signale med tkivi, celice ali organizmi. Signalno funkcijo opravljajo hormonski proteini. Celice lahko komunicirajo med seboj na daljavo s pomočjo signalnih proteinov, ki se prenašajo skozi medcelično snov.
Beljakovine imajo tudi motorično funkcijo. Vse vrste gibanja, ki so jih celice sposobne, na primer krčenje mišic, izvajajo posebni kontraktilni proteini. Beljakovine opravljajo tudi transportno funkcijo. Sposobni so pritrditi različne snovi in jih prenašati z enega mesta v celici na drugega. Na primer, krvni protein hemoglobin veže kisik in ga prenaša v vsa tkiva in organe telesa. Poleg tega imajo beljakovine tudi zaščitno funkcijo. Ko v telo vnesemo tuje beljakovine ali celice, v njem nastanejo posebne beljakovine, ki vežejo in nevtralizirajo tuje celice in snovi. In končno, energetska funkcija beljakovin je, da se s popolno razgradnjo 1 g beljakovin sprosti energija v količini 17,6 kJ.
Struktura celice.
Celica je sestavljena iz treh neločljivo povezanih delov: membrane, citoplazme in jedra, zgradba in delovanje jedra v različnih obdobjih življenja celice pa sta različni. Življenjska doba celice vključuje dve obdobji: delitev, zaradi katere nastaneta dve hčerinski celici, in obdobje med delitvami, ki se imenuje interfaza.
Celična membrana neposredno sodeluje z zunanjim okoljem in sodeluje s sosednjimi celicami. Sestavljen je iz zunanje plasti in plazemske membrane, ki se nahaja pod njo. Površinski sloj živalskih celic se imenuje glikokali. Povezuje celice z zunanjim okoljem in z vsemi snovmi, ki jih obdajajo. Njegova debelina je manjša od 1 mikrona.
Struktura celice
Celična membrana je zelo pomemben del celice. Drži skupaj vse celične komponente in razmejuje zunanje in notranje okolje.
Med celicami in zunanjim okoljem poteka stalna izmenjava snovi. Iz zunanjega okolja v celico vstopajo voda, različne soli v obliki posameznih ionov, anorganske in organske molekule. Produkti presnove, pa tudi snovi, ki se sintetizirajo v celici: beljakovine, ogljikovi hidrati, hormoni, ki nastajajo v celicah različnih žlez, se skozi membrano iz celice izločajo v zunanje okolje. Prenos snovi je ena glavnih funkcij plazemske membrane.
citoplazma- notranji poltekoči medij, v katerem potekajo glavni presnovni procesi. Nedavne študije so pokazale, da citoplazma ni nekakšna rešitev, katere komponente medsebojno delujejo v naključnih trkih. Primerjamo ga lahko z želejem, ki začne »trepetati« kot odziv na zunanje vplive. Tako citoplazma zaznava in prenaša informacije.
V citoplazmi se nahajajo jedro in različne organele, ki jih združuje v eno celoto, kar zagotavlja njihovo interakcijo in delovanje celice kot enotnega celostnega sistema. Jedro se nahaja v osrednjem delu citoplazme. Celotna notranja cona citoplazme je napolnjena z endoplazmatskim retikulumom, ki je celični organoid: sistem tubulov, veziklov in "cistern", omejenih z membranami. Endoplazmatski retikulum je vključen v presnovne procese, ki zagotavlja transport snovi iz okolja v citoplazmo in med posameznimi znotrajceličnimi strukturami, vendar je njegova glavna funkcija sodelovanje pri sintezi beljakovin, ki se izvaja v ribosomih. - mikroskopska telesa okrogle oblike s premerom 15-20 nm. Sintetizirane beljakovine se najprej kopičijo v kanalih in votlinah endoplazmatskega retikuluma in se nato transportirajo do organele in celičnih mest, kjer se porabijo.
Poleg beljakovin so v citoplazmi tudi mitohondriji, majhna telesa velikosti 0,2-7 mikronov, ki jih imenujemo "elektrarne" celic. V mitohondrijih se pojavijo redoks reakcije, ki celicam zagotavljajo energijo. Število mitohondrijev v eni celici se giblje od nekaj do nekaj tisoč.
Jedro- vitalni del celice, nadzoruje sintezo beljakovin in preko njih vse fiziološke procese v celici. V jedru celice, ki se ne deli, ločimo jedrsko membrano, jedrski sok, nukleolus in kromosome. Skozi jedrsko ovojnico poteka neprekinjena izmenjava snovi med jedrom in citoplazmo. Pod jedrno ovojnico - jedrski sok (poltekoča snov), ki vsebuje jedro in kromosome. Jedro je gosto zaobljeno telo, katerega dimenzije se lahko zelo razlikujejo, od 1 do 10 mikronov in več. Sestavljen je predvsem iz ribonukleoproteinov; sodeluje pri tvorbi ribosomov. Običajno je v celici 1-3 jedrca, včasih tudi do več sto. Jedro je sestavljeno iz RNA in beljakovin.
S prihodom celice je na Zemlji nastalo Življenje!
Se nadaljuje...


