Zapewnia związek komórki z otoczeniem. Połączenie organizmu z otoczeniem
Połączenie organizmu ze środowiskiem, z fizykochemicznego punktu widzenia, jest układem otwartym, czyli układem, w którym zachodzą procesy biochemiczne. Substancje wyjściowe pochodzą ze środowiska, a substancje, które również powstają w sposób ciągły, są usuwane. Równowaga między szybkością i stężeniem produktów reakcji wielokierunkowych w organizmie jest warunkowa, urojona, ponieważ przyjmowanie i usuwanie substancji nie kończy się. Ciągłe połączenie z otoczeniem i pozwala nam traktować żywy organizm jako system otwarty.
Słońce jest źródłem energii dla wszystkich żywych komórek. Komórki roślinne wychwytują energię światła słonecznego za pomocą chlorofilu, wykorzystując go do reakcji asymilacyjnych podczas fotosyntezy. Komórki zwierząt, grzybów, bakterii wykorzystują energię słoneczną pośrednio, dzieląc substancje organiczne syntetyzowane przez ziemską roślinę.
Część składników odżywczych komórki jest rozkładana w procesie oddychania komórkowego, dostarczając tym samym energię niezbędną do różnego rodzaju aktywności komórkowej. Proces ten odbywa się w organellach zwanych mitochondriami. Mitochondrium składa się z dwóch błon: zewnętrznej, oddzielającej organelle od cytoplazmy, oraz wewnętrznej, tworzącej liczne fałdy. Głównym produktem oddychania jest ATP. Opuszcza mitochondria i jest wykorzystywany jako źródło energii w wielu reakcjach chemicznych w cytoplazmie i błonie komórkowej. Jeśli do wykonania oddychania komórkowego potrzebny jest tlen, to oddychanie nazywa się oddychaniem tlenowym, ale jeśli reakcje zachodzą przy braku tlenu, mówi się o oddychaniu beztlenowym.
Do każdego rodzaju pracy wykonywanej w komórce energia jest wykorzystywana w jednej formie - w postaci energii z wiązań fosforanowych ATP. ATP to wysoce mobilny związek. Powstawanie ATP zachodzi w wewnętrznej błonie mitochondriów. ATP jest syntetyzowany we wszystkich komórkach podczas oddychania dzięki energii utleniania węglowodanów, tłuszczów i innych substancji organicznych. W komórkach roślin zielonych główna ilość ATP jest syntetyzowana w chloroplastach dzięki energii słonecznej. W nich podczas fotosyntezy wytwarzane jest wielokrotnie więcej ATP niż w mitochondriach. ATP rozkłada się wraz z zerwaniem wiązań fosfor-tlen i uwolnieniem energii. Dzieje się to pod wpływem enzymu ATPazy w procesie hydrolizy ATP - dodanie wody z eliminacją cząsteczki kwasu fosforowego. W rezultacie ATP jest przekształcany w ADP, a jeśli dwie cząsteczki kwasu fosforowego zostaną oddzielone, to w AMP. Reakcji rozszczepienia każdej gramocząsteczki kwasu towarzyszy uwolnienie 40 kJ. Jest to bardzo duża wydajność energetyczna, dlatego wiązania fosfor-tlen ATP są powszechnie nazywane makroergicznymi (wysokoenergetycznymi).
Wykorzystanie ATP w reakcjach wymiany plastycznej odbywa się poprzez ich sprzężenie z hydrolizą ATP. Cząsteczki różnych substancji są ładowane energią poprzez przyłączanie grupy fosforowej uwalnianej podczas hydrolizy z cząsteczki ATP, czyli poprzez fosforylację.
Cechą pochodnych fosforanowych jest to, że nie mogą opuścić komórki, chociaż ich „rozładowane” formy swobodnie przechodzą przez błonę. Dzięki temu ufosforylowane cząsteczki pozostają w komórce, dopóki nie zostaną użyte w odpowiednich reakcjach.
Odwrotny proces konwersji ADP do ATP zachodzi poprzez przyłączanie cząsteczki kwasu fosforowego do ADP, uwalnianie wody i pochłanianie dużej ilości energii.
Tak więc ATP jest uniwersalnym i bezpośrednim źródłem energii dla aktywności komórek. Tworzy to pojedynczy komórkowy zasób energii i umożliwia redystrybucję i transport z jednej części komórki do drugiej.
Transfer grupy fosforanowej odgrywa ważną rolę w reakcjach chemicznych, takich jak składanie makrocząsteczek z monomerów. Na przykład aminokwasy można łączyć w peptydy tylko wtedy, gdy zostały wcześniej ufosforylowane. Mechaniczne procesy skurczu lub ruchu, przenoszenie substancji rozpuszczonej wbrew gradientowi stężenia oraz inne procesy są związane z wydatkowaniem energii zmagazynowanej w ATP.
Proces wymiany energii można przedstawić w następujący sposób. Wysokocząsteczkowe substancje organiczne w cytoplazmie są enzymatycznie, drogą hydrolizy, przekształcane w prostsze, z których składają się: białka - w aminokwasy, poli- i disacharydy - w monosacharydy (+ glukoza), tłuszcze w glicerol i kwasy tłuszczowe. Brakuje procesów oksydacyjnych, uwalniana jest niewielka ilość energii, która nie jest wykorzystywana i przechodzi w formę termiczną. Większość komórek najpierw używa węglowodanów. Polisacharydy (skrobia roślinna i glikogen zwierzęcy) są hydrolizowane do glukozy. Utlenianie glukozy zachodzi w trzech fazach: glikolizy, dekarboksylacji oksydacyjnej (cykl Krebsa – cykl kwasu cytrynowego) i fosforylacji oksydacyjnej (łańcuch oddechowy). W cytoplazmie zachodzi glikoliza, w wyniku której jedna cząsteczka glukozy zostaje rozszczepiona na dwie cząsteczki kwasu pirogronowego z uwolnieniem dwóch cząsteczek ATP. W przypadku braku tlenu kwas pirogronowy jest przekształcany w etanol (fermentacja) lub kwas mlekowy (oddychanie beztlenowe).
Jeśli glikoliza zachodzi w komórkach zwierzęcych, sześciowęglowa cząsteczka glukozy rozpada się na dwie cząsteczki kwasu mlekowego. Ten proces jest wieloetapowy. Odbywa się to sekwencyjnie przez 13 enzymów. Podczas fermentacji alkoholowej z cząsteczki glukozy powstają dwie cząsteczki etanolu i dwie cząsteczki CO2.
Glikoliza jest fazą wspólną dla oddychania beztlenowego i tlenowego, pozostałe dwie są przeprowadzane tylko w warunkach tlenowych. Proces beztlenowego utleniania, w którym tylko część energii metabolitów jest uwalniana i wykorzystywana, jest procesem końcowym dla organizmów beztlenowych. W obecności tlenu kwas pirogronowy przechodzi do mitochondriów, gdzie w wyniku szeregu następujących po sobie reakcji zostaje całkowicie utleniony tlenowo do H2O i CO2 z jednoczesną fosforylacją ADP do ATP. Jednocześnie glikoliza daje dwie cząsteczki ATP, dwie - cykl Krebsa, 34 - łańcuch oddechowy. Wydajność netto z całkowitego utlenienia jednej cząsteczki glukozy do H2O i CO2 wynosi 38 cząsteczek.
Tak więc w organizmach tlenowych ostateczny rozkład substancji organicznych odbywa się poprzez ich utlenienie tlenem atmosferycznym do prostych substancji nieorganicznych: CO2 i H2O. Proces ten zachodzi na cristae mitochondriów. W tym przypadku uwalniana jest maksymalna ilość energii swobodnej, której znaczna część jest zarezerwowana w cząsteczkach ATP. Łatwo zauważyć, że tlenowe utlenianie w największym stopniu dostarcza komórce darmowej energii.
W wyniku katabolizmu w komórce gromadzą się bogate w energię cząsteczki ATP, a CO2 i nadmiar wody są uwalniane do środowiska zewnętrznego.
Cząsteczki cukru, które nie są potrzebne do oddychania, mogą być przechowywane w komórce. Nadmiar lipidów jest albo rozszczepiany, po czym ich produkty rozszczepienia przedostają się do mitochondriów jako substrat do oddychania, albo odkładają się w rezerwie w cytoplazmie w postaci kropelek tłuszczu. Białka zbudowane są z aminokwasów wchodzących do komórki. Synteza białek zachodzi w organellach zwanych rybosomami. Każdy rybosom składa się z dwóch podcząstek - dużej i małej: obie podcząstki zawierają cząsteczki białka i cząsteczki RNA.
Rybosomy są często przyczepione do specjalnego systemu błon, składającego się ze zbiorników i pęcherzyków, do tzw. retikulum endoplazmatycznego (ER); w komórkach wytwarzających dużo białka retikulum endoplazmatyczne jest często bardzo dobrze rozwinięte i pełne rybosomów. Niektóre enzymy są skuteczne tylko wtedy, gdy są połączone z błoną. Tutaj znajduje się większość enzymów biorących udział w syntezie lipidów. Tak więc retikulum endoplazmatyczne jest niejako rodzajem pulpitu komórkowego.
Ponadto ER dzieli cytoplazmę na oddzielne sekcje lub przedziały, tj. oddziela różne procesy chemiczne zachodzące jednocześnie w cytoplazmie, a tym samym zmniejsza prawdopodobieństwo wzajemnego zakłócania się tych procesów.
Często produkty utworzone przez daną komórkę są używane poza komórką. W takich przypadkach białka syntetyzowane na rybosomach przechodzą przez błony retikulum endoplazmatycznego i są pakowane w pęcherzyki błonowe, które tworzą się wokół nich, które są następnie wyplatane z ER. Bąbelki te, spłaszczając się i układając jeden na drugim, jak naleśniki w stosie, tworzą charakterystyczną strukturę zwaną kompleksem Golgiego lub aparatem Golgiego. Podczas pobytu w aparacie Golgiego białka ulegają pewnym zmianom. Gdy nadejdzie czas, aby opuścić komórkę, błoniaste pęcherzyki łączą się z błoną komórkową i opróżniają się, wylewając ich zawartość na zewnątrz, czyli wydzielanie następuje na drodze egzocytozy.
W aparacie Golgiego powstają również lizosomy - woreczki błonowe zawierające enzymy trawienne. Zrozumienie, w jaki sposób komórka wytwarza, pakuje i eksportuje określone białka oraz w jaki sposób „wie”, które białka powinna zachować dla siebie, jest jedną z najbardziej fascynujących gałęzi współczesnej cytologii.
Błony każdej komórki nieustannie się poruszają i zmieniają. Błony ER poruszają się powoli w całej komórce. Oddzielne sekcje tych błon są oddzielane i tworzą pęcherzyki, które tymczasowo stają się częścią aparatu Golgiego, a następnie w procesie egzocytozy łączą się z błoną komórkową.
Później materiał błony powraca do cytoplazmy, gdzie jest ponownie wykorzystywany.
Wymiana substancji, które dostają się do komórki lub są przez nią uwalniane na zewnątrz, a także wymiana różnych sygnałów z mikro- i makrośrodowiskiem, odbywa się przez zewnętrzną błonę komórki. Jak wiadomo, błona komórkowa jest dwuwarstwą lipidową, w której osadzone są różne cząsteczki białek, które działają jako wyspecjalizowane receptory, kanały jonowe, urządzenia aktywnie przenoszące lub usuwające różne chemikalia, kontakty międzykomórkowe itp. W zdrowych komórkach eukariotycznych fosfolipidy są rozmieszczone w membrana asymetrycznie: zewnętrzna powierzchnia składa się z sfingomieliny i fosfatydylocholiny, wewnętrzna powierzchnia składa się z fosfatydyloseryny i fosfatydyloetanoloaminy. Utrzymanie takiej asymetrii wymaga nakładów energetycznych. Dlatego w przypadku uszkodzenia komórki, jej infekcji, głodu energetycznego, zewnętrzna powierzchnia błony wzbogacana jest o niezwykłe dla niej fosfolipidy, które stają się sygnałem dla innych komórek i enzymów o uszkodzeniu komórki przy odpowiedniej reakcji na nią. Najważniejszą rolę odgrywa rozpuszczalna forma fosfolipazy A2, która rozkłada kwas arachidonowy i tworzy lizoformy z powyższych fosfolipidów. Kwas arachidonowy jest ogniwem ograniczającym powstawanie takich mediatorów stanu zapalnego, jak eikozanoidy, a cząsteczki ochronne - pentraksyny (białko C-reaktywne (CRP), prekursory białek amyloidowych) - przyłączane są do lizoform w błonie, a następnie aktywują układ dopełniacza wzdłuż klasycznego szlaku i niszczenia komórek.
Struktura błony przyczynia się do zachowania cech środowiska wewnętrznego komórki, jej różnic od środowiska zewnętrznego. Zapewnia to selektywna przepuszczalność błony komórkowej, istnienie w niej aktywnych mechanizmów transportowych. Ich naruszenie w wyniku bezpośredniego uszkodzenia, na przykład przez tetrodotoksynę, ouabainę, tetraetyloamon lub w przypadku niewystarczającego zaopatrzenia w energię odpowiednich „pomp”, prowadzi do naruszenia składu elektrolitów komórki, zmiany jej metabolizmu , naruszenie określonych funkcji - skurcz, przewodzenie impulsu wzbudzającego itp. Naruszenie komórkowych kanałów jonowych (wapniowych, sodowych, potasowych i chlorkowych) u ludzi może być również uwarunkowane genetycznie przez mutację genów odpowiedzialnych za budowę tych kanały. Tak zwane kanałopatie są przyczyną dziedzicznych chorób układu nerwowego, mięśniowego i pokarmowego. Nadmierne spożycie wody do wnętrza komórki może prowadzić do jej pęknięcia - cytolizy - w wyniku perforacji błony podczas aktywacji dopełniacza lub ataku limfocytów cytotoksycznych i naturalnych zabójców.
W błonie komórkowej wbudowanych jest wiele receptorów - struktur, które w połączeniu z odpowiednimi cząsteczkami sygnałowymi (ligandami) przekazują sygnał do komórki. Dzieje się to poprzez różne kaskady regulacyjne, składające się z enzymatycznie aktywnych cząsteczek, które są sekwencyjnie aktywowane i ostatecznie przyczyniają się do realizacji różnych programów komórkowych, takich jak wzrost i proliferacja, różnicowanie, ruchliwość, starzenie się i śmierć komórki. Kaskady regulacyjne są dość liczne, ale ich liczba nie została jeszcze w pełni określona. Wewnątrz komórki istnieje również system receptorów i związane z nimi kaskady regulacyjne; tworzą pewną sieć regulacyjną z punktami koncentracji, dystrybucji i wyboru dalszej ścieżki sygnału w zależności od stanu funkcjonalnego komórki, etapu jej rozwoju i jednoczesnego działania sygnałów z innych receptorów. Skutkiem tego może być zahamowanie lub wzmocnienie sygnału, jego kierunek wzdłuż innej ścieżki regulacyjnej. Zarówno aparat receptorowy, jak i szlaki transdukcji sygnału poprzez kaskady regulacyjne, np. do jądra, mogą zostać zakłócone w wyniku defektu genetycznego, który występuje jako wada wrodzona na poziomie organizmu lub w wyniku mutacji somatycznej w określonej komórce rodzaj. Mechanizmy te mogą być uszkadzane przez czynniki zakaźne, toksyny, a także zmieniać się podczas starzenia. Ostatnim etapem tego może być naruszenie funkcji komórki, procesów jej proliferacji i różnicowania.
Na powierzchni komórek znajdują się również cząsteczki, które odgrywają ważną rolę w procesach interakcji międzykomórkowych. Mogą to być białka adhezji komórek, antygeny zgodności tkankowej, antygeny specyficzne tkankowo, różnicujące itp. Zmiany w składzie tych cząsteczek powodują naruszenie oddziaływań międzykomórkowych i mogą powodować aktywację odpowiednich mechanizmów eliminacji takich komórek, ponieważ stwarzają pewne zagrożenie dla integralności organizmu jako rezerwuaru infekcji, zwłaszcza wirusowej, lub jako potencjalne inicjatory wzrostu guza.
Naruszenie dostaw energii do komórki
Źródłem energii w komórce jest pożywienie, po rozbiciu którego energia uwalniana jest do substancji końcowych. Mitochondria to główne miejsce produkcji energii, w którym substancje są utleniane za pomocą enzymów łańcucha oddechowego. Utlenianie jest głównym dostawcą energii, ponieważ w wyniku glikolizy z tej samej ilości substratów utleniania (glukozy) uwalnianych jest nie więcej niż 5% energii w porównaniu z utlenianiem. Około 60% energii uwalnianej podczas utleniania jest akumulowane przez fosforylację oksydacyjną w fosforanach makroergicznych (ATP, fosforan kreatyny), reszta jest rozpraszana w postaci ciepła. W przyszłości wysokoenergetyczne fosforany są wykorzystywane przez komórkę do takich procesów jak pompowanie, synteza, podział, ruch, sekrecja itp. Istnieją trzy mechanizmy, których uszkodzenie może spowodować zakłócenie w dostawie energii do komórki : pierwszy to mechanizm syntezy enzymów metabolizmu energetycznego, drugi to mechanizm fosforylacji oksydacyjnej, trzeci - mechanizm wykorzystania energii.
Naruszenie transportu elektronów w łańcuchu oddechowym mitochondriów lub rozdzielenie utleniania i fosforylacji ADP z utratą potencjału protonowego – siły napędowej wytwarzania ATP, prowadzi do osłabienia fosforylacji oksydacyjnej w taki sposób, że większość energii jest rozpraszana w zmniejsza się forma ciepła i liczba związków makroergicznych. Rozprzęganie utleniania i fosforylacji pod wpływem adrenaliny jest wykorzystywane przez komórki organizmów homoiotermicznych do zwiększenia produkcji ciepła przy utrzymaniu stałej temperatury ciała podczas chłodzenia lub jej podwyższeniu podczas gorączki. W tyreotoksykozie obserwuje się istotne zmiany w budowie mitochondriów i metabolizmie energetycznym. Zmiany te są początkowo odwracalne, ale po pewnym czasie stają się nieodwracalne: mitochondria ulegają fragmentacji, rozpadowi lub pęcznieniu, tracą cristae, zamieniając się w wakuole, a w końcu kumulują substancje takie jak szklistość, ferrytyna, wapń, lipofuscyna. U pacjentów ze szkorbutem mitochondria łączą się, tworząc chondrosfery, prawdopodobnie z powodu uszkodzenia błony przez związki nadtlenkowe. Znaczne uszkodzenie mitochondriów następuje pod wpływem promieniowania jonizującego, podczas transformacji normalnej komórki w złośliwą.
Mitochondria są potężnym magazynem jonów wapnia, gdzie ich stężenie jest o kilka rzędów wielkości wyższe niż w cytoplazmie. Kiedy mitochondria są uszkodzone, wapń przedostaje się do cytoplazmy, powodując aktywację proteinaz z uszkodzeniem struktur wewnątrzkomórkowych i zaburzeniem funkcji odpowiedniej komórki, na przykład przykurczami wapniowymi lub nawet „śmiercią wapniową” w neuronach. W wyniku naruszenia zdolności czynnościowych mitochondriów gwałtownie wzrasta powstawanie wolnych rodnikowych związków nadtlenkowych, które mają bardzo wysoką reaktywność i przez to uszkadzają ważne składniki komórki - kwasy nukleinowe, białka i lipidy. Zjawisko to obserwuje się podczas tzw. stresu oksydacyjnego i może mieć negatywne konsekwencje dla istnienia komórki. Tak więc uszkodzeniu zewnętrznej błony mitochondrialnej towarzyszy uwalnianie do cytoplazmy substancji zawartych w przestrzeni międzybłonowej, przede wszystkim cytochromu C i niektórych innych substancji biologicznie czynnych, które wywołują reakcje łańcuchowe powodujące zaprogramowaną śmierć komórki – apoptozę. Poprzez uszkadzanie mitochondrialnego DNA, reakcje wolnych rodników zniekształcają informację genetyczną niezbędną do tworzenia pewnych enzymów łańcucha oddechowego, które są wytwarzane specjalnie w mitochondriach. Prowadzi to do jeszcze większego zakłócenia procesów oksydacyjnych. Ogólnie rzecz biorąc, wewnętrzny aparat genetyczny mitochondriów, w porównaniu z aparatem genetycznym jądra, jest mniej chroniony przed szkodliwymi wpływami, zdolnymi do zmiany zakodowanej w nim informacji genetycznej. W efekcie dysfunkcja mitochondriów występuje przez całe życie, na przykład w procesie starzenia, podczas złośliwej transformacji komórki, a także na tle dziedzicznych chorób mitochondrialnych związanych z mutacją mitochondrialnego DNA w jaju. Obecnie opisano ponad 50 mutacji mitochondrialnych, które powodują dziedziczne choroby zwyrodnieniowe układu nerwowego i mięśniowego. Są przekazywane dziecku wyłącznie od matki, ponieważ mitochondria plemników nie są częścią zygoty, a zatem nowego organizmu.
Naruszenie ochrony i przekazywania informacji genetycznej
Jądro komórkowe zawiera większość informacji genetycznej, a tym samym zapewnia jej normalne funkcjonowanie. Za pomocą selektywnej ekspresji genów koordynuje pracę komórki w interfazie, przechowuje informację genetyczną, odtwarza i przekazuje materiał genetyczny w procesie podziału komórki. Replikacja DNA i transkrypcja RNA zachodzą w jądrze. Różne czynniki chorobotwórcze, takie jak promieniowanie ultrafioletowe i jonizujące, utlenianie wolnych rodników, chemikalia, wirusy, mogą uszkadzać DNA. Szacuje się, że każda komórka zwierzęcia stałocieplnego w ciągu 1 dnia. traci ponad 10 000 baz. Do tego należy dodać naruszenia podczas kopiowania podczas dzielenia. Gdyby to uszkodzenie trwało, komórka nie byłaby w stanie przetrwać. Ochrona polega na istnieniu potężnych systemów naprawczych, takich jak endonukleaza ultrafioletowa, replikacja naprawcza i rekombinacyjne systemy naprawcze, które zastępują uszkodzenia DNA. Defekty genetyczne w układach naprawczych powodują rozwój chorób ze względu na zwiększoną wrażliwość na czynniki uszkadzające DNA. Jest to kseroderma barwnikowa, a także niektóre zespoły przyspieszonego starzenia, którym towarzyszy zwiększona skłonność do występowania nowotworów złośliwych.
System regulacji procesów replikacji DNA, transkrypcji informacyjnego RNA (mRNA), translacji informacji genetycznej z kwasów nukleinowych na strukturę białek jest dość złożony i wielopoziomowy. Oprócz kaskad regulacyjnych, które wyzwalają działanie ponad 3000 czynników transkrypcyjnych, które aktywują określone geny, istnieje również wielopoziomowy system regulacyjny, w którym pośredniczą małe cząsteczki RNA (interferujące RNA; RNAi). Genom ludzki, który składa się z około 3 miliardów zasad purynowych i pirymidynowych, zawiera tylko 2% genów strukturalnych odpowiedzialnych za syntezę białek. Pozostałe zapewniają syntezę regulatorowych RNA, które wraz z czynnikami transkrypcyjnymi aktywują lub blokują pracę genów strukturalnych na poziomie DNA w chromosomach lub wpływają na translację informacyjnego RNA (mRNA) podczas tworzenia cząsteczki polipeptydu w cytoplazmie . Naruszenie informacji genetycznej może wystąpić zarówno na poziomie genów strukturalnych, jak i regulacyjnej części DNA z odpowiednimi objawami w postaci różnych chorób dziedzicznych.
Ostatnio wiele uwagi poświęca się zmianom w materiale genetycznym, które zachodzą podczas indywidualnego rozwoju organizmu i wiążą się z hamowaniem lub aktywacją niektórych odcinków DNA i chromosomów poprzez ich metylację, acetylację i fosforylację. Zmiany te utrzymują się przez długi czas, niekiedy przez całe życie organizmu od embriogenezy do starości i nazywane są dziedziczeniem epigenomicznym.
Rozmnażaniu komórek ze zmienioną informacją genetyczną zapobiegają również systemy (czynniki) kontroli cyklu mitotycznego. Oddziałują z cyklinozależnymi kinazami białkowymi i ich podjednostkami katalitycznymi - cyklinami - i blokują przejście przez komórkę pełnego cyklu mitotycznego, zatrzymując podział na granicy między fazą presyntetyczną i syntetyczną (blok G1/S) do czasu zakończenia naprawy DNA , a jeśli to niemożliwe, inicjują komórki zaprogramowanej śmierci. Czynniki te obejmują gen p53, którego mutacja powoduje utratę kontroli nad proliferacją transformowanych komórek; występuje w prawie 50% ludzkich nowotworów. Drugi punkt kontrolny przejścia cyklu mitotycznego znajduje się na granicy G2/M. Tutaj prawidłowe rozmieszczenie materiału chromosomalnego między komórkami potomnymi w mitozie lub mejozie jest kontrolowane za pomocą kompleksu mechanizmów kontrolujących wrzeciono komórki, centrum i centromery (kinetochory). Nieskuteczność tych mechanizmów prowadzi do naruszenia rozkładu chromosomów lub ich części, co objawia się brakiem dowolnego chromosomu w jednej z komórek potomnych (aneuploidia), obecnością dodatkowego chromosomu (poliploidalność), oddzieleniem część chromosomu (delecja) i jej przeniesienie do innego chromosomu (translokacja) . Takie procesy są bardzo często obserwowane podczas rozmnażania złośliwie zdegenerowanych i transformowanych komórek. Jeśli dzieje się to podczas mejozy z komórkami rozrodczymi, prowadzi albo do śmierci płodu we wczesnym stadium rozwoju embrionalnego, albo do narodzin organizmu z chorobą chromosomową.
Niekontrolowane rozmnażanie komórek podczas wzrostu guza następuje w wyniku mutacji w genach kontrolujących proliferację komórek i są one nazywane onkogenami. Wśród ponad 70 obecnie znanych onkogenów większość z nich jest składnikami regulacji wzrostu komórek, niektóre to czynniki transkrypcyjne regulujące aktywność genów, a także czynniki hamujące podział i wzrost komórek. Kolejnym czynnikiem ograniczającym nadmierną ekspansję (rozprzestrzenianie się) proliferujących komórek jest skracanie końców chromosomów – telomerów, które w wyniku oddziaływania czysto sterycznego nie są w stanie się w pełni replikować, dlatego po każdym podziale komórki telomery są skracane o pewna część baz. Tak więc proliferujące komórki dorosłego organizmu, po określonej liczbie podziałów (zwykle od 20 do 100, w zależności od typu organizmu i jego wieku), wyczerpują długość telomerów i dalsza replikacja chromosomów ustaje. Zjawisko to nie występuje w nabłonku spermatogennym, enterocytach i komórkach embrionalnych ze względu na obecność enzymu telomerazy, który przywraca długość telomerów po każdym podziale. W większości komórek organizmów dorosłych telomeraza jest zablokowana, ale niestety jest aktywowana w komórkach nowotworowych.
Połączenie między jądrem a cytoplazmą, transport substancji w obu kierunkach odbywa się przez pory w błonie jądrowej przy udziale specjalnych systemów transportowych o zużyciu energii. W ten sposób energia i substancje plastyczne, cząsteczki sygnałowe (czynniki transkrypcyjne) są transportowane do jądra. Odwrócony przepływ wprowadza do cytoplazmy cząsteczki mRNA i transferowego RNA (tRNA), rybosomy niezbędne do syntezy białek w komórce. Ten sam sposób transportu substancji jest nieodłączny od wirusów, w szczególności takich jak HIV. Przenoszą swój materiał genetyczny do jądra komórki gospodarza z jego dalszym włączeniem do genomu gospodarza i przeniesieniem nowo utworzonego wirusowego RNA do cytoplazmy w celu dalszej syntezy białek nowych cząstek wirusa.
Naruszenie procesów syntezy
Procesy syntezy białek zachodzą w cysternach retikulum endoplazmatycznego, ściśle związanego z porami błony jądrowej, przez które rybosomy, tRNA i mRNA przedostają się do retikulum endoplazmatycznego. Tutaj przeprowadzana jest synteza łańcuchów polipeptydowych, które później uzyskują ostateczną formę w agranularnej retikulum endoplazmatycznym i kompleksie blaszkowym (kompleks Golgiego), gdzie ulegają modyfikacji potranslacyjnej i asocjacji z cząsteczkami węglowodanów i lipidów. Nowo powstałe cząsteczki białka nie pozostają w miejscu syntezy, ale za pomocą złożonego regulowanego procesu, który nazywa się kineza białkowa, są aktywnie przenoszone do tej izolowanej części komórki, w której będą pełnić zamierzoną funkcję. W tym przypadku bardzo ważnym krokiem jest ustrukturyzowanie przenoszonej cząsteczki w odpowiednią konfigurację przestrzenną, zdolną do pełnienia swojej wrodzonej funkcji. Taka struktura zachodzi za pomocą specjalnych enzymów lub na matrycy wyspecjalizowanych cząsteczek białka - chaperonów, które pomagają nowo powstałej lub zmienionej pod wpływem czynników zewnętrznych cząsteczce białka uzyskać prawidłową trójwymiarową strukturę. W przypadku niekorzystnego wpływu na komórkę, gdy istnieje możliwość naruszenia struktury cząsteczek białka (na przykład ze wzrostem temperatury ciała, procesem zakaźnym, zatruciem), stężenie białek opiekuńczych w komórce gwałtownie wzrasta. Dlatego takie cząsteczki są również nazywane białka stresu, lub białka szoku cieplnego. Naruszenie struktury cząsteczki białka prowadzi do powstania chemicznie obojętnych konglomeratów, które w przypadku amyloidozy, choroby Alzheimera itp. odkładają się w komórce lub poza nią. w tym przypadku, jeśli struktura pierwotna nie zaszła prawidłowo, wszystkie kolejne cząsteczki również będą wadliwe. Taka sytuacja ma miejsce w tzw. chorobach prionowych (scrappie u owiec, wścieklizna u krów, kuru, choroba Creutzfeldta-Jakoba u ludzi), gdy defekt jednego z białek błonowych komórki nerwowej powoduje późniejsze nagromadzenie mas obojętnych wewnątrz komórka i zakłócenie jej życiowej aktywności.
Naruszenie procesów syntezy w komórce może nastąpić na różnych jej etapach: transkrypcja RNA w jądrze, translacja polipeptydów w rybosomach, modyfikacja potranslacyjna, hipermetylacja i glikozylacja beżowej cząsteczki, transport i dystrybucja białek w komórce oraz ich usuwanie na zewnątrz. W tym przypadku można zaobserwować wzrost lub spadek liczby rybosomów, rozpad polirybosomów, rozszerzenie cystern ziarnistej retikulum endoplazmatycznego, utratę przez nie rybosomów, tworzenie pęcherzyków i wakuoli. Tak więc w przypadku zatrucia bladym muchomorem enzym polimerazy RNA zostaje uszkodzony, co zakłóca transkrypcję. Toksyna błonicza, inaktywując czynnik wydłużenia, zaburza procesy translacji, powodując uszkodzenie mięśnia sercowego. Przyczyną naruszenia syntezy niektórych określonych cząsteczek białka mogą być czynniki zakaźne. Na przykład herpeswirusy hamują syntezę i ekspresję cząsteczek antygenu MHC, co pozwala im częściowo uniknąć kontroli immunologicznej, a pałeczki dżumy hamują syntezę mediatorów ostrego zapalenia. Pojawienie się nietypowych białek może zatrzymać ich dalszy rozkład i doprowadzić do nagromadzenia obojętnego lub nawet toksycznego materiału. W pewnym stopniu może się do tego przyczynić również zakłócenie procesów rozpadu.
Naruszenie procesów rozpadu
Równolegle z syntezą białka w komórce następuje jego rozpad. W normalnych warunkach ma to istotne znaczenie regulacyjne i formacyjne, na przykład podczas aktywacji nieaktywnych form enzymów, hormonów białkowych i białek cyklu mitotycznego. Prawidłowy wzrost i rozwój komórek wymaga dokładnie kontrolowanej równowagi między syntezą a degradacją białek i organelli. Jednak w procesie syntezy białek, z powodu błędów w działaniu aparatu syntetyzującego, nieprawidłowej struktury cząsteczki białka, jej uszkodzenia przez czynniki chemiczne i bakteryjne, stale powstaje dość duża liczba wadliwych cząsteczek. Według niektórych szacunków ich udział wynosi około jednej trzeciej wszystkich syntetyzowanych białek.
Komórki ssaków mają kilka głównych szlaki degradacji białek: poprzez proteazy lizosomalne (hydrolazy pentydowe), proteinazy zależne od wapnia (endopeptydazy) i układ proteasomów. Ponadto istnieją również wyspecjalizowane proteinazy, takie jak kaspazy. Głównym organellem, w którym zachodzi rozkład substancji w komórkach eukariotycznych, jest lizosom, który zawiera liczne enzymy hydrolityczne. W wyniku procesów endocytozy i różnego rodzaju autofagii w lizosomach i fagolizosomach niszczeniu ulegają zarówno wadliwe cząsteczki białek, jak i całe organelle: uszkodzone mitochondria, fragmenty błony komórkowej, niektóre białka zewnątrzkomórkowe, zawartość ziarnistości wydzielniczych.
Ważnym mechanizmem degradacji białka jest proteasom, złożona multikatalityczna struktura proteinazy zlokalizowana w cytozolu, jądrze, retikulum endoplazmatycznym i na błonie komórkowej. Ten układ enzymatyczny jest odpowiedzialny za rozkład uszkodzonych białek, a także zdrowych białek, które muszą zostać usunięte, aby komórki mogły normalnie funkcjonować. W tym przypadku niszczone białka łączy się wstępnie ze specyficznym polipeptydem ubikwityny. Jednak nieubikwitowane białka mogą być również częściowo zniszczone w proteasomach. Rozpad cząsteczki białka w proteasomach na krótkie polipeptydy (przetwarzanie) z późniejszą prezentacją wraz z cząsteczkami MHC typu I jest ważnym ogniwem we wdrażaniu immunologicznej kontroli homeostazy antygenowej organizmu. W przypadku osłabienia funkcji proteasomu dochodzi do akumulacji uszkodzonych i niepotrzebnych białek, co towarzyszy starzeniu się komórek. Naruszenie degradacji białek zależnych od cyklin prowadzi do naruszenia podziału komórek, degradacji białek wydzielniczych - do rozwoju mukowiscydozy. Odwrotnie, wzrost funkcji proteasomów towarzyszy wyczerpaniu organizmu (AIDS, rak).
W przypadku genetycznie uwarunkowanych naruszeń degradacji białek organizm jest niezdolny do życia i umiera we wczesnych stadiach embriogenezy. Jeśli rozkład tłuszczów lub węglowodanów jest zaburzony, pojawiają się choroby akumulacyjne (tezaurismozy). Jednocześnie w komórce gromadzi się nadmierna ilość niektórych substancji lub produktów ich niepełnego rozkładu - lipidów, polisacharydów, co znacznie zaburza funkcję komórki. Najczęściej obserwuje się ją w nabłonkach wątroby (hepatocytach), neuronach, fibroblastach i makrofagocytach.
Nabyte zaburzenia w procesach rozpadu substancji mogą wystąpić w wyniku procesów patologicznych (na przykład dystrofii białka, tłuszczu, węglowodanów i pigmentu) i towarzyszyć im tworzenie się nietypowych substancji. Naruszenia w układzie proteolizy lizosomalnej prowadzą do zmniejszenia adaptacji podczas głodu lub zwiększonego obciążenia, do wystąpienia niektórych dysfunkcji endokrynologicznych - obniżenia poziomu insuliny, tyreoglobuliny, cytokin i ich receptorów. Naruszenia degradacji białek spowalniają tempo gojenia się ran, powodują rozwój miażdżycy i wpływają na odpowiedź immunologiczną. W warunkach niedotlenienia, zmian wewnątrzkomórkowego pH, uszkodzenia popromiennego, charakteryzującego się zwiększoną peroksydacją lipidów błonowych, a także pod wpływem substancji lizosomotropowych - endotoksyn bakteryjnych, metabolitów grzybów toksycznych (sporofusarin), kryształów tlenku krzemu - stabilność błony lizosomów zmiany, aktywowane enzymy lizosomalne są uwalniane do cytoplazmy, co powoduje zniszczenie struktur komórkowych i ich śmierć.
Rozdział 1
PODSTAWY FIZJOLOGII KOMÓREK
I. Dudel
błona plazmatyczna . Komórki zwierzęce są ograniczone przez błonę plazmatyczną (ryc. 1.1). Bardziej szczegółowo zajmiemy się jego strukturą, która jest bardzo podobna do budowy wielu błon wewnątrzkomórkowych. Główna matryca membrany składa się z lipidy głównie fosfatydylocholina. Te lipidy składają się z hydrofilowej grupy czołowej, do której przyłączone są długie hydrofobowe łańcuchy węglowodorowe. W wodzie takie lipidy spontanicznie tworzą dwuwarstwowy film o grubości 4-5 nm, w którym hydrofilowe grupy są skierowane do ośrodka wodnego, a hydrofobowe łańcuchy węglowodorowe są ułożone w dwóch rzędach, tworząc bezwodną fazę lipidową. Błony komórkowe są dwuwarstwami lipidowymi tego typu i zawierają glikolipidy, cholesterol i fosfolipidy (ryc. 1.2). Część hydrofilową glikolipidów tworzą oligosacharydy. Glikolipidy zawsze znajdują się na zewnętrznej powierzchni błony komórkowej, a część oligosacharydowa cząsteczki jest zorientowana jak włos zanurzony w środowisku. Rozproszone wśród fosfolipidów w prawie równych ilościach cząsteczki cholesterolu stabilizują błonę. Rozkład różnych lipidów w wewnętrznej i zewnętrznej warstwie błony nie jest taki sam, a nawet w tej samej warstwie występują obszary, w których skoncentrowane są określone typy lipidów. Taki nierówny rozkład
Ryż. 1,1. Schematyczny rysunek komórki przedstawiający najważniejsze organelle
prawdopodobnie ma pewne, dotychczas niejasne znaczenie funkcjonalne.
Głównymi elementami funkcjonalnymi zanurzonymi we względnie obojętnej matrycy lipidowej błony są wiewiórki(Rys. 1.2). Waga białka waha się od 25 do 75% w różnych błonach, ale ponieważ cząsteczki białka są znacznie większe niż cząsteczki lipidów, 50% wagowo odpowiada stosunkowi 1 cząsteczki białka do 50 cząsteczek lipidu. Niektóre białka przenikają przez błonę od jej zewnętrznej do wewnętrznej powierzchni, podczas gdy inne są utrwalane w jednej warstwie. Cząsteczki białka są zwykle zorientowane tak, że ich grupy hydrofobowe są zanurzone w błonie lipidowej, a polarne grupy hydrofilowe na powierzchni błony są zanurzone w fazie wodnej. Wiele białek błony zewnętrznej powierzchni to glikoproteiny; ich hydrofilowe grupy sacharydowe stykają się ze środowiskiem pozakomórkowym.
Systemy błonowe organelli wewnątrzkomórkowych .
Około połowę objętości komórki zajmują organelle izolowane z cytozolu przez błony. Całkowita powierzchnia błon organelli wewnątrzkomórkowych jest co najmniej 10 razy większa od powierzchni błony plazmatycznej. Najczęściej stosowanym systemem membranowym jest retikulum endoplazmatyczne, reprezentujący sieć

Ryż. 1.2.Schematyczne przedstawienie błony plazmatycznej. Białka są osadzone w dwuwarstwie fosfolipidowej, przy czym niektóre obejmują dwuwarstwę, podczas gdy inne są zakotwiczone tylko w warstwie zewnętrznej lub wewnętrznej
bardzo skręcone kanaliki lub wydłużone struktury torebkowe; duże obszary retikulum endoplazmatycznego są usiane rybosomami; taka siateczka nazywana jest ziarnistą lub szorstką (ryc. 1.1). Aparat Golgiego składa się również z blaszek związanych z błoną, z których odrywają się pęcherzyki (ryc. 1.1). Lizosomy i peroksysomy są małymi wyspecjalizowanymi pęcherzykami. We wszystkich tych różnorodnych organellach błona i przestrzeń, którą obejmuje, zawierają określone zestawy enzymów; wewnątrz organelli gromadzą się specjalne produkty przemiany materii, które są wykorzystywane do wykonywania różnych funkcji organelli.
RdzeńI mitochondria różnią się tym, że każda z tych organelli jest otoczona dwiema błonami. Jądro odpowiada za kinetyczną kontrolę metabolizmu; pofałdowana wewnętrzna błona mitochondrialna jest miejscem metabolizmu oksydacyjnego; tutaj, w wyniku utleniania pirogronianu lub kwasów tłuszczowych, syntetyzowany jest wysokoenergetyczny związek adenozynotrifosforan (ATP lub ATP).
cytoszkielet . Cytoplazmy otaczającej organelle w żadnym wypadku nie można uznać za amorficzną; jest przesiąknięta siecią cytoszkieletu. Cytoszkielet składa się z mikrotubul, filamentów aktynowych i filamentów pośrednich (ryc. 1.1). mikrotubule mieć średnicę zewnętrzną około 25 nm; powstają, jak normalny polimer, w wyniku połączenia cząsteczek białka tubuliny. włókna aktynowe- włókna kurczliwe znajdujące się w warstwie przybłonowej oraz w całej komórce – biorą udział głównie w procesach związanych z ruchem. Włókna pośrednie składają się z bloków o różnym składzie chemicznym w różnych typach komórek; tworzą różnorodne powiązania między pozostałymi dwoma elementami cytoszkieletu, o których mowa powyżej. Organelle i błona komórkowa są również związane z cytoszkieletem, który nie tylko zachowuje kształt komórki i położenie w niej organelli, ale także determinuje zmianę kształtu komórki i jej ruchliwość.
Cytozol . Około połowę objętości komórki zajmuje cytozol. Ponieważ zawiera około 20% (wagowo) białka, jest bardziej żelem niż roztworem wodnym. Małe cząsteczki, w tym organiczne i nieorganiczne jony, rozpuszczony w fazie wodnej. Między komórką a środowiskiem (przestrzeń zewnątrzkomórkowa) następuje wymiana jonów; te procesy wymiany zostaną omówione w następnej sekcji. Stężenie jonów w przestrzeni pozakomórkowej utrzymuje się z dużą dokładnością na stałym poziomie; wewnątrzkomórkowe stężenie każdego z jonów ma również określony poziom, który różni się od tego na zewnątrz komórki (tabela 1.1). Najczęstszym kationem w środowisku pozakomórkowym jest Na+ w komórce jego stężenie jest ponad 10 razy mniejsze. Wręcz przeciwnie, wewnątrz komórki stężenie K + jest najwyższe, poza komórką jest niższe o ponad rząd wielkości. Największy gradient między stężeniami pozakomórkowymi i wewnątrzkomórkowymi występuje dla Ca 2+, którego stężenie wolnych jonów wewnątrz komórki jest co najmniej 10 000 razy mniejsze niż na zewnątrz. Nie wszystkie jony są rozpuszczone w cytozolu, część z nich jest adsorbowana na białkach lub odkładana w organellach. Na przykład w przypadku Ca 2+ jony związane są znacznie liczniejsze niż wolne. Większość białek cytozolu to enzymy, przy udziale których odbywa się wiele procesów metabolizmu pośredniego: glikoliza i glukoneogeneza, synteza lub niszczenie aminokwasów, synteza białek na rybosomach (ryc. 1.1). Cytozol zawiera również kropelki lipidów i granulki glikogenu, które służą jako rezerwy ważnych cząsteczek.
Tabela 1.1.Wewnątrz- i zewnątrzkomórkowe stężenia jonów w komórkach mięśniowych zwierząt homoiotermicznych. ALE – – „aniony komórkowe o wysokiej masie cząsteczkowej”
|
Stężenie wewnątrzkomórkowe |
Stężenie pozakomórkowe |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
Z l - |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO 3 - |
8 mmol/l |
HCO 3 - |
27 mmol/l |
|
A- |
155 mmol/l |
Inny kationy |
5 mmol/l |
|
Potencjał spoczynkowy -90 mV |
|||
1.2. Wymiana substancji między komórką a środowiskiem
Opisaliśmy pokrótce strukturę komórki, aby wykorzystać ten opis do przeglądu podstaw fizjologii komórki. W żadnym przypadku komórka nie może być uważana za formację statyczną, ponieważ między różnymi przedziałami wewnątrzkomórkowymi, a także między przedziałami a środowiskiem zachodzi stała wymiana substancji. Struktury komórki znajdują się w dynamicznej równowadze, a wzajemne oddziaływanie komórek ze sobą oraz ze środowiskiem zewnętrznym jest niezbędnym warunkiem utrzymania życia funkcjonującego organizmu. W tym rozdziale rozważymy podstawowe mechanizmy takiej wymiany. W dalszych rozdziałach mechanizmy te zostaną omówione w odniesieniu do komórki nerwowej i jej funkcji;
jednak te same mechanizmy leżą u podstaw funkcjonowania wszystkich innych narządów.
Dyfuzja.Najprostszym procesem przenoszenia substancji jest dyfuzja. W roztworach (lub gazach) atomy i cząsteczki poruszają się swobodnie, a różnica stężeń jest równoważona przez dyfuzję. Rozważ dwie objętości wypełnione cieczą lub gazem (ryc. 1.3), w których substancje mają stężenia c1 i c2 i oddzielone warstwą o polu powierzchni A i grubości D. Przepływ materii m w czasie t opisane Pierwsze prawo dyfuzji Ficka:
dm/ dt= DA/ D ( C 1 –С 2)=DA/ DD C(1)
gdzie D jest współczynnikiem dyfuzji, który jest stały dla danej substancji, rozpuszczalnika i temperatury. W bardziej ogólnej formie dla różnicy stężeń dc w odległości dx
dm/dt= -D A dc/dx,(2)
przepływ przez odcinek A jest proporcjonalny do gradientu stężenia dc/dx . W równaniu pojawia się znak minus, ponieważ zmiana stężenia w kierunku x jest ujemna.
Dyfuzja to najważniejszy proces, w którym większość cząsteczek w roztworach wodnych przemieszcza się na krótkie odległości. Dotyczy to również ich ruchu w komórce, o ile dyfuzja nie jest utrudniana przez błony. Wiele substancji może swobodnie dyfundować przez błony lipidowe, zwłaszcza woda i rozpuszczone gazy, takie jak O 2 i CO 2 . rozpuszczalny w tłuszczach

Ryż. 1.3.Ilościowy schemat dyfuzji. Dwie przestrzenie są oddzielone warstwą grubościDi obszar ALE. C; - wysokie stężenie cząstek w lewej części objętości, C:, - niskie stężenie cząstek w prawej części części, różowa powierzchnia to gradient stężenia w warstwie dyfuzyjnej. Strumień dyfuzji dm/dt – patrz równanie (1)
substancje również dobrze dyfundują przez błony; dotyczy to również stosunkowo małych cząsteczek polarnych, takich jak etanol i mocznik, podczas gdy cukry z trudem przechodzą przez warstwę lipidową. Jednocześnie warstwy lipidowe są praktycznie nieprzepuszczalne dla naładowanych cząsteczek, w tym nawet jonów nieorganicznych. W przypadku nieelektrolitów równanie dyfuzji (1) jest zwykle przekształcane przez połączenie właściwości membrany i substancji dyfundującej w jedno przepuszczalność parametrów (P):
dm/dt=P AD C.(3)
Na ryc. 1.4 w porównaniu przepuszczalność (P) błony lipidowej dla różnych cząsteczek.
Dyfuzja przez pory membrany . Błona plazmatyczna (i inne błony komórkowe) są przepuszczalne nie tylko dla substancji dyfundujących przez warstwę lipidową, ale także dla wielu jonów, cukrów, aminokwasów i nukleotydów. Substancje te przechodzą przez membranę przez pory utworzone przez białka transportowe, osadzony w membranie. Wewnątrz takich białek znajduje się wypełniony wodą kanał o średnicy poniżej 1 nm, przez który mogą dyfundować małe cząsteczki. Poruszają się po gradiencie stężeń, a jeśli przenoszą ładunek, to ich ruch przez kanały jest również regulowany przez potencjał błonowy. Kanały membranowe są stosunkowo selektywne

Ryż. 1.4.Przepuszczalność sztucznych dwuwarstw lipidowych dla różnych substancji
w zależności od rodzaju molekuł, które mogą przez nie przejść, istnieją np. kanały potasowe, sodowe i wapniowe, z których każdy jest nieprzepuszczalny dla prawie każdego jonu poza konkretnym. Taki selektywność ze względu na ładunek lub strukturę miejsc wiążących w ściankach kanału, co ułatwia transport określonej cząsteczki i zapobiega przenikaniu innych substancji przez kanał (ryc. 1,5, A) .
Za zachowaniem membranowe kanały jonoweŁatwo to zaobserwować, ponieważ prąd powstający podczas ruchu jonów można zmierzyć i to nawet dla pojedynczego kanału. Pokazano, że kanały spontanicznie iz dużą częstotliwością zmieniają swój stan z otwartego na zamknięty. Kanał potasowy charakteryzuje się impulsami prądowymi o amplitudzie około 2 pA (2 10-12 A) i czasie trwania kilku milisekund (patrz ryc. 2.12, s. 37) [3]. W tym okresie przechodzą przez nią dziesiątki tysięcy jonów. Przejście białek z jednej konformacji do drugiej jest badane za pomocą dyfrakcji rentgenowskiej, spektroskopii Mössbauera i magnetycznego rezonansu jądrowego (NMR). Białka są zatem wysoce dynamicznymi, ruchomymi strukturami, a kanał przez białko to nie tylko sztywna, wypełniona wodą rurka (ryc. 1.5A), ale labirynt szybko poruszających się grup molekularnych i ładunków. Ta dynamiczna odpowiedź kanału znajduje odzwierciedlenie w profil energetyczny kanału, pokazano na ryc. 1,5, B. Tutaj odcięta pokazuje długość kanału od roztworu zewnętrznego o stężeniu jonów C0 i potencjale 0 do roztworu wewnętrznego o stężeniu C1 i potencjale E. Oś y

Ryż. 1.5.A. Schemat białka tworzącego kanał potasowy osadzony w podwójnej warstwie lipidowej błony komórkowej. Cztery ładunki ujemne są zamocowane na „ścianie” kanału. B. Schemat profilu energetycznego kanału pokazanego na ryc. A. Na osi y wykreślane są wartości energii kinetycznej niezbędnej do przejścia kanału; wzdłuż osi odciętej odległości między wewnętrzną i zewnętrzną powierzchnią błony. Minima energetyczne odpowiadają miejscom wiązania dodatnio naładowanych jonów o stałych ładunkach ujemnych w ściance kanału. Maksima energii odpowiadają przeszkodom dyfuzyjnym w kanale. Zakłada się, że konformacja białka kanału samorzutnie oscyluje; opcje profili energetycznych są pokazane liniami ciągłymi i przerywanymi; oscylacje te znacznie ułatwiają wiązanie jonów przy pokonywaniu bariery energetycznej (ale ze zmianami)
pokazano poziomy energii jonów w miejscach wiązania kanałów; szczyt na wykresie reprezentuje barierę przepuszczalności, którą energia jonów musi pokonać, aby przeniknąć do kanału, a „dołek” na wykresie reprezentuje względnie stabilny stan (wiązanie). Pomimo niedrożności szczytu energii, jon może przeniknąć kanał, jeśli profil energii spontanicznie się zmienia; w ten sposób jon może nagle pojawić się „po drugiej stronie” szczytu energii i może dalej przemieszczać się do komórki. W zależności od ładunku, wielkości i stopnia uwodnienia jonu oraz jego zdolności do wiązania się ze strukturami ścianek kanału, profil energetyczny kanału jest różny dla różnych jonów, co może wyjaśniać selektywność poszczególnych typów kanałów.
Równowaga dyfuzji jonów . Dyfuzja różnych jonów przez kanały błonowe powinna prowadzić do eliminacji różnic stężeń między środowiskami zewnątrz- i wewnątrzkomórkowymi. Jednak, jak widać z tabeli. 1.1 takie różnice się utrzymują, więc muszą być jakieś równowaga między dyfuzją a innymi procesami transportu przez błonę. Następne dwie sekcje dotyczą sposobów, w jakie taka równowaga jest ustalana. W przypadku jonów na równowagę dyfuzji wpływa ich ładunek. Dyfuzję nienaładowanych cząsteczek zapewnia różnica stężeń dc , a gdy stężenia się wyrównają, faktyczny transport zostaje zatrzymany. Naładowane cząstki są dodatkowo pod wpływem pola elektrycznego. Na przykład, gdy jon potasu opuszcza komórkę wzdłuż gradientu stężenia, przenosi jeden ładunek dodatni. W ten sposób środowisko wewnątrzkomórkowe staje się bardziej ujemnie naładowane, co powoduje różnicę potencjałów w poprzek błony. Wewnątrzkomórkowy ładunek ujemny zapobiega opuszczaniu komórki przez nowe jony potasu, a te jony, które mimo to opuszczają komórkę, dodatkowo zwiększają ładunek na błonie. Przepływ jonów potasu zatrzymuje się, gdy działanie pola elektrycznego kompensuje ciśnienie dyfuzji z powodu różnicy stężeń. Jony nadal przechodzą przez błonę, ale w równych ilościach w obu kierunkach. Dlatego dla danej różnicy stężeń jonów na membranie istnieje potencjał równowagi mi jon w którym zatrzymuje się przepływ jonów przez membranę. Potencjał równowagi można łatwo określić za pomocą Równania Nernsta:
mijon= rT/ zF* jaNa zewnątrz/ C w(4)
gdzie R to stała gazowa, T to temperatura bezwzględna, z to wartościowość jonów (ujemna dla anionów) Na zewnątrz jest stężenie jonów pozakomórkowych, C w jest wewnątrzkomórkowe stężenie jonu, F Numer Faradaya. Jeśli do równania podstawimy stałe, to w temperaturze ciała (T = 310 K) potencjał równowagi dla jonów potasu E K równa się:
Ek= –61 mDziennik B /(5)
Jeśli [ K + out ]/[ K + in ] = 39, jak wynika z tabeli. 1.1, więc
Ek= -61 m B log 39= -97 mV.
Rzeczywiście stwierdzono, że wszystkie komórki mają: potencjał błonowy; w komórkach mięśniowych ssaków jego poziom wynosi około -90 mV. W zależności od warunków i względnych stężeń jonów, komórki mogą mieć potencjał błonowy w zakresie od -40 do -120mV. Dla komórki w powyższym przykładzie (Tabela 1.1) potencjał spoczynkowy, równy około -90 mV, wskazuje, że strumienie jonów potasu przez kanały błony są w przybliżeniu w równowadze. Nie jest to zaskakujące, ponieważ najbardziej prawdopodobny jest stan otwarty kanałów potasowych w błonie spoczynkowej; membrana jest najbardziej przepuszczalna dla jonów potasu. Potencjał błonowy jest jednak determinowany również przez strumienie innych jonów.
Łatwość, z jaką nienaładowane cząstki mogą dyfundować przez membranę, określa się ilościowo w równaniu (3). Przepuszczalność dla naładowanych cząstek opisany nieco bardziej złożonym równaniem:
P= m rT/ dF(6)
gdzie mto ruchliwość jonu w błonie, D – grubość membrany, a R , T i F znane stałe termodynamiczne. Wyznaczone w ten sposób wartości przepuszczalności dla różnych jonów można wykorzystać do obliczenia potencjału błonowego Em gdy jony potasu, sodu i chloru przechodzą jednocześnie przez membranę (z przepuszczalnością P K, P Na i P Cl odpowiednio). Zakłada się, że potencjał w membranie spada równomiernie, dzięki czemu natężenie pola jest stałe. W tym przypadku ma zastosowanie Równanie Goldmana lub równanie pola stałego :
mim= R T/ F * ln(PK + P Na + P Cl )/ (PK + P Na + P Cl )(7)
Dla większości błon komórkowych P K około 30 razy wyższa niż R Na (patrz także sekcja 1.3). Wartość względna PCl różni się znacznie; dla wielu membran PCl mały w porównaniu do R K , jednak dla innych (np. w mięśniach szkieletowych) PCl , znacznie wyższy niż R K.
Aktywny transport, pompa sodowa . W poprzedniej części opisano bierną dyfuzję jonów i wynikający z niej potencjał błonowy przy danych stężeniach jonów wewnątrz- i zewnątrzkomórkowych. Jednak w wyniku tego procesu stężenie jonów wewnątrz komórki nie jest automatycznie stabilizowane, ponieważ membrana
potencjał jest nieco bardziej elektroujemny niż E K , i znacznie więcej w porównaniu do E Na (około +60 mV). Ze względu na dyfuzję wewnątrzkomórkowe stężenia jonów, przynajmniej potasu i sodu, powinny wyrównać się z zewnątrzkomórkowymi. Stabilność gradientu jonów osiąga się dzięki aktywnemu transportowi: białka błonowe transportują jony przez błonę wbrew gradientom elektrycznym i (lub) stężeń, zużywając do tego energię metaboliczną. Najważniejszym procesem aktywnego transportu jest praca Na/K - pompa, która istnieje w prawie wszystkich komórkach;
pompa wypompowuje jony sodu z komórki, jednocześnie pompując jony potasu do komórki. Zapewnia to niskie wewnątrzkomórkowe stężenie jonów sodu i wysokie potasu (tab. 1.1). Gradient stężenia jonów sodu na błonie pełni określone funkcje związane z przekazywaniem informacji w postaci impulsów elektrycznych (patrz rozdział 2.2), a także z utrzymywaniem innych aktywnych mechanizmów transportu i regulacji objętości komórek (patrz niżej). Dlatego nie dziwi fakt, że ponad 1/3 energii zużywanej przez ogniwo jest wydatkowana na pompę Na/K, a w niektórych najbardziej aktywnych ogniwach aż do 70% energii zużywa się na jej działanie.
Białko transportowe Na/K jest ATPazą. Na wewnętrznej powierzchni membrany rozkłada ATP na ADP i fosforan (ryc. 1.6). Energia jednej cząsteczki ATP jest wykorzystywana do transportu trzech jonów sodu z komórki i jednocześnie dwóch jonów potasu do komórki, czyli w sumie jeden ładunek dodatni jest usuwany z komórki w jednym cyklu. Zatem pompa Na/K jest elektrogeniczny(wytwarza prąd elektryczny przez błonę), co prowadzi do wzrostu elektroujemności potencjału błony o około 10 mV. Białko transportowe wykonuje tę operację z dużą szybkością: od 150 do 600 jonów sodu na sekundę. Sekwencja aminokwasowa białka transportowego jest znana, ale mechanizm tego złożonego transportu wymiennego nie jest jeszcze jasny. Proces ten opisano za pomocą profili energetycznych przenoszenia jonów sodu lub potasu przez białka (rys. 1.5.5). Ze względu na charakter zmiany tych profili, związanej z ciągłymi zmianami konformacji białka transportowego (proces wymagający energii), można ocenić stechiometrię wymiany: dwa jony potasu są wymieniane na trzy jony sodu.
Pompa Na/K, jak izolowana Na+ ATP-aza błonowa zależna od /K+, specyficznie hamowana przez glikozydową ouabainę sercową (strofantynę). Ponieważ praca pompy Na/K jest wieloetapową reakcją chemiczną, tak jak wszystkie reakcje chemiczne, jest ona w dużej mierze zależna od temperatury, co

Ryż. 1.6.Schemat pompy-ATPazy Na/K (zanurzonej w podwójnej warstwie lipidowej błony komórkowej), która w jednym cyklu wyprowadza z komórki trzy jony Na+ wbrew gradientom potencjału i stężenia oraz wprowadza do komórki dwa jony K + . Podczas tego procesu jedna cząsteczka ATP zostaje podzielona na ADP i fosforan. Na schemacie ATPaza jest pokazana jako dimer składający się z dużej (funkcjonalnej) i małej podjednostki; w błonie występuje jako tetramer złożony z dwóch dużych i dwóch małych podjednostek
pokazano na ryc. 1.7. Tutaj przepływ jonów sodu z komórek mięśniowych jest pokazany w funkcji czasu; jest to praktycznie równoważne przepływowi jonów sodu za pośrednictwem pompy Na/K, ponieważ pasywny przepływ jonów sodu w stosunku do gradientów stężeń i potencjałów jest niezwykle mały. Jeżeli preparat zostanie schłodzony o ok. 18°C, to przepływ jonów sodu z ogniwa szybko zmniejszy się 15-krotnie i zaraz po podgrzaniu powróci do pierwotnego poziomu. Taki spadek przepływu jonów sodu z ogniwa jest kilkakrotnie większy niż ten, który odpowiadałby zależności temperaturowej procesu dyfuzji lub prostej reakcji chemicznej. Podobny efekt obserwuje się, gdy energia metaboliczna zostaje wyczerpana w wyniku zatrucia dinitrofenolem (DNP) (ryc. 1.7.5). Dlatego wypływ jonów sodu z komórki zapewnia reakcja zależna od energii – aktywna pompa. Inną cechą pompy, obok znacznej zależności od temperatury i energii, jest obecność poziomu nasycenia (jak w przypadku wszystkich innych reakcji chemicznych); oznacza to, że prędkość pompy nie może wzrastać w nieskończoność wraz ze wzrostem stężenia transportowanych jonów (rys. 1.8). Natomiast przepływ substancji biernie dyfundującej rośnie proporcjonalnie do różnicy stężeń zgodnie z prawem dyfuzji (równania 1 i 2).

Ryż. 1.7. A, B. Na aktywny transport + . Oś Y: przepływ radioaktywnego 24 Na + z ogniwa (imp./min). oś odciętych: czas od rozpoczęcia eksperymentu. ALE. Ogniwo jest schładzane od 18,3°C do 0,5°C; pływ Na+ poza komórką w tym okresie jest zahamowany. B. Tłumienie wypływu Na+ z ogniwa dinitrofenolem (DNF) w stężeniu 0,2 mmol/l (z późniejszymi zmianami)
Oprócz pompy Na/K membrana plazmowa zawiera co najmniej jeszcze jedną pompę - wapń; pompa ta wypompowuje jony wapnia (Ca 2+) z komórki i bierze udział w utrzymywaniu ich wewnątrzkomórkowego stężenia na skrajnie niskim poziomie (tabela 1.1). Pompa wapniowa jest obecna w bardzo dużej gęstości w siateczce sarkoplazmatycznej komórek mięśniowych, które gromadzą jony wapnia w wyniku rozpadu cząsteczek ATP (patrz rozdział 4).
Wpływ pompy Na/K na potencjał błonowy i objętość komórek . Na ryc. 1.9 pokazuje różne składniki prądu błonowego i pokazuje wewnątrzkomórkowe stężenia jonów, które

Ryż. 1.8.Stosunek szybkości transportu cząsteczek do ich stężenia (w miejscu wejścia do kanału lub w miejscu przyłączenia pompy) podczas dyfuzji przez kanał lub podczas transportu pompującego. Ten ostatni nasyca się w wysokich stężeniach (maksymalna szybkość, Vmaks ) wartość na odciętej odpowiadająca połowie maksymalnej prędkości pompy ( Vmaks /2), jest stężeniem równowagowym DO m

Ryż. 1.9.Schemat przedstawiający stężenia Na+ , K+ i Cl- wewnątrz i na zewnątrz komórki oraz w jaki sposób jony te przenikają przez błonę komórkową (poprzez określone kanały jonowe lub za pomocą pompy Na/K. Przy danych gradientach stężeń potencjały równowagi E Na, E K i E C l - są równe wskazanym, potencjał błonowy Em = – 90 mV
zapewnić ich istnienie. Zewnętrzny prąd jonów potasu obserwuje się przez kanały potasowe, ponieważ potencjał błonowy jest nieco bardziej elektrododatni niż potencjał równowagi jonów potasu. Całkowite przewodnictwo kanałów sodowych jest znacznie niższe niż kanałów potasowych; przy potencjale spoczynkowym kanały sodowe są otwarte znacznie rzadziej niż kanały potasowe; jednak mniej więcej taka sama liczba jonów sodu wchodzi do komórki, jak jony potasu ją opuszczają, ponieważ do dyfuzji jonów sodu do komórki potrzebne są duże stężenia i gradienty potencjału. Pompa Na/K zapewnia idealną kompensację pasywnych prądów dyfuzyjnych, ponieważ transportuje jony sodu z komórki i jony potasu do niej. Tym samym pompa jest elektrogeniczna ze względu na różnicę w ilości ładunków przenoszonych do iz ogniwa, co przy normalnej szybkości jej działania tworzy potencjał błonowy około 10 mV jest bardziej elektroujemny niż gdyby był generowany przez same pasywne prądy jonowe (patrz równanie 7). W rezultacie potencjał błonowy zbliża się do potencjału równowagi potasu, co zmniejsza wyciek jonów potasu. Na aktywność Pompa /K jest regulowana przez wewnątrzkomórkowe stężenie jonów sodu. W miarę zmniejszania się stężenia jonów sodu do usunięcia z komórki prędkość pompy zmniejsza się (rys. 1.8), dzięki czemu praca pompy i przepływ jonów sodu do komórki równoważą się, utrzymując wewnątrzkomórkowe stężenie jonów sodu na poziomie około 10 mmol/l.
Aby utrzymać równowagę między pompowaniem a pasywnymi prądami membranowymi, potrzeba o wiele więcej cząsteczek Na/K niż jest białek kanałowych dla jonów potasu i sodu. Gdy kanał jest otwarty, w ciągu kilku milisekund przechodzą przez niego dziesiątki tysięcy jonów (patrz wyżej), a ponieważ kanał jest zwykle otwierany kilka razy na sekundę, w tym czasie przechodzi przez niego łącznie ponad 105 jonów. Pojedyncze białko pompujące porusza kilkaset jonów sodu na sekundę, więc błona plazmatyczna musi zawierać około 1000 razy więcej cząsteczek pompujących niż cząsteczki kanałowe. Pomiary prądów kanałowych w spoczynku wykazały średnio jeden otwarty kanał potasowy i jeden sodowy na membranę 1 µm 2; wynika z tego, że w tej samej przestrzeni powinno znajdować się około 1000 cząsteczek pompy Na/K, tj. odległość między nimi wynosi średnio 34 nm; średnica białka pompującego jako białka kanałowego wynosi 8–10 nm. W ten sposób membrana jest wystarczająco gęsto nasycona cząsteczkami pompującymi
Fakt, że przepływ jonów sodu do komórki, a jonów potasu z komórki jest kompensowany pracą pompy, ma jeszcze jedną konsekwencję, polegającą na utrzymywaniu stabilnego ciśnienia osmotycznego i stała objętość. Wewnątrz komórki występuje duże stężenie dużych anionów, głównie białek (A – w tabeli 1.1), które nie są w stanie przeniknąć przez błonę (lub przenikać bardzo wolno) i dlatego są stałym składnikiem wewnątrz komórki. Aby zrównoważyć ładunek tych anionów, potrzebna jest taka sama liczba kationów. Ze względu na działanie pompy Na/K kationy te to głównie jony potasu. Istotny wzrost wewnątrzkomórkowego stężenia jonów mógł nastąpić dopiero przy wzroście stężenia anionów na skutek przepływu C1 - wzdłuż gradientu stężenia do komórki (tab. 1.1), ale przeciwdziała temu potencjał błonowy. Prąd przychodzący Cl- obserwowane tylko do osiągnięcia potencjału równowagi dla jonów chlorkowych; obserwuje się to, gdy gradient jonów chlorkowych jest prawie przeciwny do gradientu jonów potasowych, ponieważ jony chlorkowe są naładowane ujemnie (równanie 4). W ten sposób ustala się niskie wewnątrzkomórkowe stężenie jonów chlorkowych, odpowiadające niskiemu zewnątrzkomórkowemu stężeniu jonów potasu. Rezultatem jest ograniczenie całkowitej liczby jonów w komórce. Jeżeli potencjał błonowy spada, gdy pompa Na/K jest zablokowana, na przykład podczas niedotlenienia, wówczas potencjał równowagi dla jonów chlorkowych spada, a wewnątrzkomórkowe stężenie jonów chlorkowych odpowiednio wzrasta. Przywracając równowagę ładunków, jony potasu również dostają się do komórki; wzrasta całkowite stężenie jonów w komórce, co zwiększa ciśnienie osmotyczne; to zmusza wodę do wejścia do komórki. Komórka puchnie. Ten obrzęk jest widoczny in vivo w warunkach braku energii.
gradient stężenia Na + jako siła napędowa transportu membranowego . Znaczenie pompy Na/K– dla komórki nie ogranicza się do stabilizacji normalnych gradientów K+ i Na+ w poprzek błony. Energia zmagazynowana w gradiencie membrany Na+ , jest często używany do zapewnienia transportu membranowego dla innych substancji. Na przykład na ryc. 1.10 pokazuje "symport" Na+ i cząsteczki cukru do komórki. Białko transportujące przez błonę przenosi cząsteczkę cukru do komórki nawet wbrew gradientowi stężenia, podczas gdy Na + porusza się wzdłuż gradientu stężenia i potencjału, dostarczając energię do transport cukrów. Taki transport cukrów zależy całkowicie od istnienia wysokiego gradientu Na+ ; jeśli stężenie wewnątrzkomórkowe Na+ znacznie wzrasta, wtedy transport cukrów ustaje. Dla różnych C Acharow, istnieją różne systemy symportów. Transport aminokwasów w klatce podobnej do transportu C akharov pokazany na ryc. 1.10; zapewnia to również gradient Na+ , istnieje co najmniej pięć różnych systemów symportów, z których każdy specjalizuje się w jednej grupie pokrewnych aminokwasów.
Oprócz systemów symportowych istnieją również „antyport”. Jeden z nich na przykład przenosi jeden jon wapnia z komórki w jednym cyklu w zamian za trzy przychodzące jony sodu (ryc. 1.10). Energia dla transport Ca 2+ powstaje w wyniku wejścia trzech jonów sodu wzdłuż gradientu stężenia i potencjału. Ta energia jest wystarczająca (na potencjale spoczynkowym) do utrzymania wysokiego gradientu jonów wapniowych (od mniej niż 10–7 mol/l wewnątrz komórki do około 2 mmol/l na zewnątrz komórki).
Endo- i egzocytoza . W przypadku niektórych substancji, które dostają się do komórki lub muszą zostać usunięte

Ryż. 1.10.Białka osadzone w podwójnej warstwie lipidowej błony pośredniczą w imporcie glukozy i Na+ do komórki, a także Ca2+/Na+ -antyport, w którym siłą napędową jest gradient Na+ na błonie komórkowej
z niego nie ma kanałów transportowych; takie substancje obejmują na przykład białka i cholesterol. Mogą przejść przez błonę plazmatyczną do pęcherzyki lub bąbelki, przez endo- i egzocytozę. Na ryc. 1.11 pokazuje główne mechanizmy tych procesów. Podczas egzocytozy pewne organelle (patrz niżej) tworzą pęcherzyki wypełnione substancją, którą należy usunąć z komórki, taką jak hormony lub enzymy zewnątrzkomórkowe. Kiedy takie pęcherzyki docierają do błony komórkowej, ich błona lipidowa łączy się z nią, umożliwiając w ten sposób ucieczkę zawartości do środowiska zewnętrznego. W odwrotnym procesie, endocytozie, błona plazmatyczna wnika, tworząc jamę, która następnie pogłębia się i zamyka, tworząc wewnątrzkomórkowy pęcherzyk wypełniony płynem zewnątrzkomórkowym i niektórymi makrocząsteczkami. Aby zapewnić połączenie błon i zamknięcie pęcherzyka, elementy kurczliwe cytoszkieletu działają w połączeniu z samymi błonami (patrz poniżej). Endocytoza nie zawsze polega na prostym wychwyceniu ośrodka zewnątrzkomórkowego do komórki. Błona komórkowa zawiera, często zorganizowane w wyspecjalizowane grupy, specyficzne receptory dla makrocząsteczek, takich jak insulina czy antygeny. Po związaniu tych makrocząsteczek z ich receptorami dochodzi do endocytozy w obszarze błony otaczającej receptor, a makrocząsteczka jest selektywnie transportowana do komórki (ryc. 1.12, B).
Endo- i egzocytoza występują w komórkach w sposób ciągły. Ilość krążącego materiału membranowego jest znaczna; w ciągu 1 godziny makrofag wchłania w postaci pęcherzyków dwukrotnie większą powierzchnię swojej błony cytoplazmatycznej. W większości komórek obrót materiału błonowego nie jest tak intensywny, ale nadal powinien być znaczny.

Ryż. 1.11.egzocytoza i endocytoza. W górę: pęcherzyk wewnątrzkomórkowy łączy się z dwuwarstwą lipidową błony komórkowej i otwiera się do przestrzeni zewnątrzkomórkowej. Ten proces nazywa się egzocytozą. Na dnie: błona plazmatyczna wdziera się na niewielkim obszarze i oddziela pęcherzyk wypełniony materiałem pozakomórkowym. Ten proces nazywa się endocytozą.
1.3. Transport substancji w komórce
Endo- i egzocytoza to nie tylko procesy transportu substancji przez błonę komórkową, ale także procesy wymiany błonowej - strukturalnych składników samej komórki. Przedmiotem rozważań w tej części są inne podobne procesy transportowe w komórce i jej organellach.

Ryż. 1.12. A-B. Schemat procesów, w tym egzo- i endocytoza. ALE. Białko syntetyzowane w ziarnistym retikulum endoplazmatycznym jest transportowane aparatem Golgiego do błony plazmatycznej, gdzie jest wydzielane na drodze egzocytozy. B. Cholesterol związany z cząsteczkami LDL (lipoproteiny o niskiej gęstości) przyłącza się do błony komórkowej, indukuje tworzenie się pęcherzyka endocytowego w tym obszarze błony i jest transportowany do lizosomów, gdzie jest uwalniany. W. Materiał zewnątrzkomórkowy wychwycony podczas endocytozy (na rycinie po prawej), transportowany przez komórkę w pęcherzykach lub pęcherzykach i uwalniany przez egzocytozę (na rycinie lewy)
Dyfuzja . Oczywiście w cytozolu różnica stężeń jest eliminowana przez dyfuzję; to samo dotyczy płynów zawartych w organellach. Ze względu na wysokie stężenie rozpuszczonego białka dyfuzja jest tu znacznie wolniejsza niż w wodzie. Błony lipidowe — wokół komórki i wewnątrz organelli — to dwuwymiarowe płyny, w których zachodzi dyfuzja. Lipidy w dwuwarstwowej błonie dyfundują we własnej warstwie, rzadko przechodząc od jednej do drugiej. Zanurzone w nich białka są również dość mobilne; obracają się wokół osi prostopadłej do błony lub dyfundują bocznie z bardzo różnymi stałymi dyfuzji, 2–10 000 razy wolniej niż fosfolipidy. Jeśli więc niektóre białka poruszają się swobodnie w warstwie lipidowej i z taką samą prędkością, jak same cząsteczki lipidów, to inne są zakotwiczone, tj. dość silnie związany z cytoszkieletem. W błonie znajdują się „stałe” agregaty określonych białek, takie jak pre- i postsynaptyczne struktury komórek nerwowych. Swobodnie poruszające się białka można zademonstrować poprzez wiązanie ich z barwnikami fluorescencyjnymi, które są indukowane do świecenia poprzez krótkie oświetlenie niewielkiego obszaru błony krótkimi błyskami. Takie eksperymenty pokazują, że w czasie krótszym niż 1 minuta białka związane z barwnikiem są równomiernie rozłożone na membranie w odległości do 10 μm.
Aktywny transport w błonach organelli .
Procesy aktywnego transportu, które odgrywają istotną rolę w funkcjonowaniu błony komórkowej, zachodzą również wewnątrz komórki, w błonach organelli. Specyficzna zawartość różnych organelli powstaje częściowo w wyniku syntezy wewnętrznej, a częściowo w wyniku aktywnego transportu z cytozolu. Jednym z przykładów tego ostatniego jest wspomniana powyżej pompa Ca 2+ w siateczce sarkoplazmatycznej komórek mięśniowych. Szczególnie interesujące jest to, że w przypadku syntezy ATP w mitochondriach odwrotna zasada dotyczy tego, co zachodzi w pompach ATPazy błony komórkowej (ryc. 1.6). Podczas syntezy ATP metabolizm oksydacyjny prowadzi do powstania stromego gradientu H+ na błonach wewnętrznych. Gradient ten jest siłą napędową procesu odwrotnego do cyklu pompowania aktywnego transportu molekularnego: jony H+ przemieszczają się przez błonę wzdłuż gradientu, a uwolniona w wyniku tego energia zapewnia syntezę ATP z ADP i fosforanu . Powstały ATP z kolei dostarcza energię do komórki, w tym do aktywnego transportu.
Transport w pęcherzykach . Komórka ma dużą liczbę organelli i związanych z nimi pęcherzyków (ryc. 1.1). Organelle te, a zwłaszcza pęcherzyki, są w ciągłym ruchu, przenosząc swoją zawartość do innych organelli lub do błony komórkowej. Pęcherzyki mogą również migrować z błony komórkowej do organelli, tak jak w endocytozie.
Proces sekrecja białka pokazano na ryc. 1.12 ALE. Białko jest syntetyzowane w pobliżu jądra komórkowego na rybosomach związanych z retikulum endoplazmatycznym (tzw. retikulum endoplazmatyczne ziarniste lub szorstkie); W retikulum endoplazmatycznym białko jest pakowane do pęcherzyków transportowych, które są oddzielane od organelli i migrują do aparatu Golgiego. Tutaj łączą się ze zbiornikami aparatu Golgiego, gdzie białko jest modyfikowane (tj. Przekształcane w glikoproteinę). Na końcach cystern pęcherzyki ponownie się rozdzielają. Pęcherzyki wydzielnicze niosące zmodyfikowane białko przemieszczają się w kierunku błony komórkowej i uwalniają swoją zawartość na drodze egzocytozy.
Inny przykład ścieżki transportu w komórce pokazano na ryc. 1.12, B; jest pobieranie cholesterolu przez komórkę. Cholesterol transportowany we krwi jest związany głównie z białkami, takimi jak cząsteczki „lipoproteina o niskiej gęstości”(LNP). Cząsteczki te przyczepiają się do określonych miejsc receptora LDL na błonie, gdzie zachodzi endocytoza, a LDL jest transportowany do komórki w „powlekanych” pęcherzykach. Pęcherzyki te łączą się, tworząc endosomy i tracą „pierścienie” podczas tego procesu. Endosomy z kolei łączą się z pierwotnymi lizosomami zawierającymi głównie enzymy hydrolityczne, tworząc drugorzędowe, większe lizosomy. W nich cholesterol uwalniany jest z cząsteczek LDL i dyfunduje do cytozolu, gdzie staje się dostępny np. do syntezy błon lipidowych. Pęcherzyki niezawierające LDL są również oddzielane od endosomów, które w szczególny sposób przemieszczają się do błony komórkowej i łączą się z nią, zwracając materiał błony i prawdopodobnie receptory LDL. Od momentu związania cząsteczki LDL z błoną mija 10–15 minut, aż cholesterol zostanie uwolniony z wtórnego lizosomu. Zaburzenia w wiązaniu i wchłanianiu LDL, czyli w dostarczaniu cholesterolu do komórki, odgrywają decydującą rolę w rozwoju poważnej i rozległej choroby miażdżycy („stwardnienie” tętnic).
Istnieje wiele innych tras transportowych podobnych do tych pokazanych na ryc. 1.11 i 1.12,A, za pomocą których w komórce poruszają się określone pęcherzyki. Nie wiadomo dokładnie, w jaki sposób się poruszają, ale prawdopodobnie w ten proces biorą udział elementy cytoszkieletu. Pęcherzyki mogą ślizgać się wzdłuż mikrotubul, w którym to przypadku energia do ruchu wydaje się być dostarczana przez białko związane z pęcherzykami, ATPazę (patrz poniżej). Zupełnie niezrozumiałe pozostaje, ile różnych pęcherzyków, poruszając się jeden po drugim we wszystkich kierunkach, dociera do celu. Oczywiście muszą być „oznaczone” w taki sposób, aby było to rozpoznawane przez system transportowy i zamieniane na celowy ruch.
Transport przez tworzenie i niszczenie organelli . Do tej pory uważaliśmy endo- i egzocytozę za procesy transportu zawartości pęcherzyków. Istnieje inny aspekt tych procesów, który polega na tym, że ukierunkowane usunięcie błony komórkowej w jednym obszarze powierzchni komórki przez endocytozę i przeciwnie, jej dodanie do drugiego przez egzocytozę, przesuwa znaczne odcinki błona (ryc. 1.12.E), dając komórce możliwość, na przykład, uformowania wyrostka lub ruchu.
Podobne przegrupowania są również typowe dla cytoszkieletu, zwłaszcza mikrofilamentów i mikrotubul (ryc. 1.1). Mikrofilamenty składają się głównie z białko F-aktyny który jest zdolny do składania się w wiązki włókniste w wyniku polimeryzacji monomeru z cytozolu. Wiązki są spolaryzowane, tj. często wyrastają tylko z jednego końca, gromadząc nowe cząsteczki aktyny, podczas gdy drugi koniec jest obojętny lub następuje tu rozłożenie. Dzięki temu spolaryzowanemu wzrostowi mikrofilamenty poruszają się sprawnie, a struktura ich sieci może się zmieniać. Przejście aktyny ze stanu zdepolimeryzowanego (zol) do stanu zorganizowanego (żel) może nastąpić bardzo szybko pod wpływem innych białek lub zmian stężenia jonów (patrz niżej). Istnieją również białka, które powodują rozpad włókien aktynowych na krótkie fragmenty. Cienkie wyrostki wielu komórek - filopodia - zawierają centralną wiązkę aktyny (ryc. 1.1), a różne ruchy filopodiów są prawdopodobnie spowodowane przejściami aktyny: polimeryzacja - depolimeryzacja.
mikrotubuleczęsto również przechodzą podobne ruchy. Mechanizm tych ruchów jest podobny - polimeryzacja tubuliny z cytozolu w taki sposób, że jeden z końców mikrotubuli rośnie, a drugi albo się nie zmienia, albo następuje w nim demontaż. W ten sposób mikrotubula, poprzez odpowiednie dodanie lub usunięcie materiału, może przemieszczać się przez cytozol.
Aktywne ruchy cytoszkieletu . Zmiany w strukturach cytoszkieletu mogą wystąpić w wyniku zarówno aktywnych ruchów, jak i opisanych powyżej przegrupowań. W wielu przypadkach ruch mikrotubul i włókien aktynowych jest napędzany przez białka kurczliwe, które wiążą włókna lub kanaliki i mogą przemieszczać je względem siebie. Wiewiórki miozyna i dyneina obecny w cytozolu wszystkich komórek w stosunkowo wysokich stężeniach; są to elementy, które przekształcają energię w ruch w wyspecjalizowanych komórkach (mięśnie) i organellach (rzęski). W komórkach mięśniowych miozyna tworzy grube włókna zorientowane równolegle do włókien aktynowych. Cząsteczka miozyny swoją „głową” przyłącza się do włókna aktynowego i wykorzystując energię ATP, wypiera miozynę wzdłuż cząsteczki aktyny. Miozyna następnie odłącza się od aktyny. Zestaw wielu takich cykli łączenia-rozłączania prowadzi do makroskopowego skurcz włókien mięśniowych(rozdz. 4). Dyneina odgrywa podobną rolę w ruchu mikrotubul podczas operacji rzęsek (ryc. 1.1). W cytoplazmie niewyspecjalizowanych komórek miozyna i dyneina nie tworzą regularnych włókien, ale w większości przypadków małe grupy cząsteczek. Nawet w postaci tak małych agregatów są w stanie poruszać włóknami aktynowymi lub mikrotubulami. Ryż. 1.13 ilustruje ten proces, gdy przeciwnie spolaryzowane cząsteczki miozyny są również przyłączone do dwóch włókien aktynowych spolaryzowanych w przeciwnych kierunkach. Główki miozyny wyginają się w kierunku ogona cząsteczki, zużywając ATP, a dwa filamenty aktynowe przesuwają się w przeciwnym kierunku, po czym miozyna odrywa się od nich. Tego rodzaju ruchy, podczas których energia ATP zamieniana jest na pracę mechaniczną, mogą zmieniać kształt cytoszkieletu, a co za tym idzie komórek, a także zapewniać transport organelli związanych z cytoszkieletem.
Procesy transportu wewnątrzkomórkowego można najdobitniej zademonstrować na aksonach komórki nerwowej. transport aksonów omówiono tutaj szczegółowo, aby zilustrować zdarzenia, które prawdopodobnie występują w podobny sposób w większości komórek. Akson, który ma zaledwie kilka mikronów średnicy, może mieć metr lub więcej, a przemieszczenie białek na drodze dyfuzji z jądra do dystalnego końca aksonu zajęłoby lata. Od dawna wiadomo, że kiedy jakakolwiek część aksonu ulega zwężeniu, proksymalna część aksonu rozszerza się. Wygląda na to, że przepływ odśrodkowy jest zablokowany w aksonie. Taki przepływ-szybki transport aksonów może pokazać ruch znaczników radioaktywnych, jak w eksperymencie pokazanym na ryc. 1.14. Radioaktywnie znakowaną leucynę wstrzyknięto do zwoju korzenia grzbietowego, a następnie, od 2 do 10 godziny, mierzono radioaktywność w nerwie kulszowym w odległości 166 mm od ciał neuronów. Przez 10 godzin szczyt radioaktywności w miejscu wstrzyknięcia nie zmienił się znacząco. Jednak fala radioaktywności rozchodziła się wzdłuż aksonu ze stałą prędkością około 34 mm na 2 godziny, czyli 410 mm/dzień. Wykazano, że we wszystkich neuronach zwierząt homoiotermicznych szybki transport aksonów zachodzi w jednakowym tempie i nie ma zauważalnych różnic między cienkimi, niezmielinizowanymi włóknami a najgrubszymi aksonami, a także między włóknami motorycznymi i czuciowymi. Rodzaj znacznika promieniotwórczego również nie wpływa na szybkość szybkiego transportu aksonów; Jako markery mogą służyć różne substancje radioaktywne.

Ryż. 1.13.Niemięśniowy kompleks miozyny o określonej orientacji może wiązać się z włóknami aktynowymi o różnej polarności i, wykorzystując energię ATP, przesuwać je względem siebie.
cząsteczki, takie jak różne aminokwasy, które są wbudowywane w białka organizmu neuronu. Jeśli przeanalizujemy obwodową część nerwu w celu określenia charakteru transportowanych tu nośników radioaktywności, to nośniki takie występują głównie we frakcji białkowej, ale także w składzie mediatorów i wolnych aminokwasów. Wiedząc, że właściwości tych substancji są różne, a zwłaszcza rozmiary ich cząsteczek, możemy wyjaśnić stałą szybkość transportu jedynie mechanizmem transportu wspólnym dla nich.
Opisane powyżej szybki transport aksonów jest wsteczny tj. skierowane z dala od ciała komórki. Wykazano, że niektóre substancje przemieszczają się z obwodu do ciała komórki za pomocą transport wsteczny. Na przykład acetylocholinoesteraza jest transportowana w tym kierunku z szybkością dwa razy mniejszą niż szybkość szybkiego transportu aksonów. Marker często używany w neuroanatomii – peroksydaza chrzanowa – również porusza się wstecznie. Transport wsteczny prawdopodobnie odgrywa ważną rolę w regulacji syntezy białek w ciele komórki. Kilka dni po przecięciu aksonów w ciele komórki obserwuje się chromatolizę, co wskazuje na naruszenie syntezy białek. Czas wymagany do chromatolizy koreluje z czasem trwania transportu wstecznego z miejsca przecięcia aksonu do ciała komórki. Taki wynik sugeruje również wyjaśnienie tego naruszenia – zakłócona zostaje transmisja z obrzeża „substancji sygnałowej”, która reguluje syntezę białek. Oczywiście główne „pojazdy” używane do szybkich aksonów
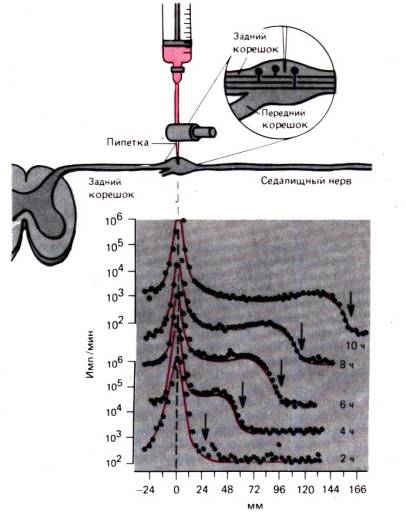
Ryż. 1.14.Eksperyment demonstrujący szybki transport aksonów we włóknach czuciowych nerwu kulszowego kota. Leucyna znakowana trytem jest wstrzykiwana do zwoju korzenia grzbietowego i radioaktywność w zwoju i włóknach czuciowych jest mierzona 2, 4, 6, 8 i 10 godzin po wstrzyknięciu. (dół rysunku). Przez odcięta odległość od zwoju do odcinków nerwu kulszowego, gdzie dokonywany jest pomiar, jest odroczona. Na osi y, tylko dla górnej i dolnej krzywych, radioaktywność (imp./min) wykreślono w skali logarytmicznej. „Fala” zwiększonej radioaktywności (strzałki) porusza się z prędkością 410mm/dzień (o )
transport są pęcherzyki (pęcherzyki) i organelle, takie jak transportowane substancje zawierające mitochondria. Ruch największych pęcherzyków lub mitochondriów można obserwować pod mikroskopem in vivo . Cząsteczki takie wykonują krótkie, szybkie ruchy w jednym kierunku, zatrzymują się, często poruszają się lekko do tyłu lub w bok, zatrzymują się ponownie, a następnie pędzą w głównym kierunku. 410 mm/dzień odpowiada średniej prędkości postępowej około 5 μm/s; prędkość każdego pojedynczego ruchu powinna być zatem znacznie wyższa, a jeśli weźmiemy pod uwagę wielkość organelli, włókien i mikrotubul, to ruchy te są naprawdę bardzo szybkie. Szybki transport aksonów wymaga znacznego stężenia ATP. Trucizny, takie jak niszcząca mikrotubule kolchicyna, również blokują szybki transport aksonów. Wynika z tego, że w rozważanym przez nas procesie transportu pęcherzyki i organelle poruszają się wzdłuż mikrotubul i włókien aktynowych; ruch ten zapewniają małe agregaty cząsteczek dyneiny i miozyny działające jak pokazano na ryc. 1.13, wykorzystując energię ATP.
Szybki transport aksonów może być również zaangażowany w: procesy patologiczne. Niektóre wirusy neurotropowe (na przykład wirusy opryszczki lub polio) wnikają do aksonu na obwodzie i przemieszczają się za pomocą transportu wstecznego do ciała neuronu, gdzie namnażają się i wywierają działanie toksyczne. Toksyna tężcowa, białko wytwarzane przez bakterie, które wnika do organizmu poprzez zmiany skórne, jest wychwytywane przez zakończenia nerwowe i transportowane do ciała neuronu, gdzie powoduje charakterystyczne skurcze mięśni. Znane są przypadki toksycznego wpływu na sam transport aksonów, np. narażenie na rozpuszczalnik przemysłowy akrylamid. Ponadto uważa się, że patogeneza beri-beri beri-beri i polineuropatii alkoholowej obejmuje naruszenie szybkiego transportu aksonów.
Oprócz szybkiego transportu aksonów w komórce występuje również dość intensywny powolny transport aksonów. Tubulina porusza się wzdłuż aksonu z szybkością około 1 mm/dzień, podczas gdy aktyna porusza się szybciej, do 5 mm/dzień. Inne białka również migrują z tymi składnikami cytoszkieletu; na przykład wydaje się, że enzymy są związane z aktyną lub tubuliną. Szybkości ruchu tubuliny i aktyny są z grubsza zgodne z szybkością wzrostu stwierdzoną dla mechanizmu opisanego wcześniej, gdy cząsteczki są włączane do aktywnego końca mikrotubuli lub mikrofilamentu. Dlatego ten mechanizm może leżeć u podstaw powolnego transportu aksonów. Szybkość powolnego transportu aksonów również w przybliżeniu odpowiada szybkości wzrostu aksonów, co najwyraźniej wskazuje na ograniczenia nałożone przez strukturę cytoszkieletu na drugi proces.
Kończąc ten rozdział, należy podkreślić, że komórki nie są w żadnym wypadku strukturami statycznymi, jak to widać na przykład na fotografiach z mikroskopu elektronowego. błona plazmatyczna a szczególnie organelle są w ciągłym szybkim ruchu i ciągłej restrukturyzacji; to jedyny powód, dla którego są w stanie funkcjonować. Co więcej, nie są to proste komory, w których zachodzą reakcje chemiczne, ale wysoko zorganizowane konglomeraty membran i włókien, w którym reakcje przebiegają w optymalnie zorganizowanej kolejności.
1.4. Regulacja funkcji komórkowych
Utrzymanie pojedynczej komórki jako jednostki funkcjonalnej jest w dużej mierze regulowane przez jądro; badanie takich mechanizmów regulacyjnych jest przedmiotem biologii i biochemii komórki. Jednocześnie komórki muszą modyfikować swoje funkcje zgodnie z warunkami środowiskowymi i potrzebami innych komórek organizmu, czyli służą jako obiekty regulacji funkcjonalnej. Poniżej krótko rozważymy, jak te regulacyjne wpływy działają na błonę plazmatyczną i jak docierają do organelli wewnątrzkomórkowych.
Wpływ regulacyjny na błonę komórkową
Potencjał błonowy . W wielu przypadkach regulacja funkcji komórkowych odbywa się poprzez zmianę potencjału błonowego. Lokalne zmiany potencjału są możliwe, gdy: 1) przez błonę przepływa prąd z sąsiedniego obszaru komórki lub generowany przez inną komórkę; 2) zmienia się zewnątrzkomórkowe stężenie jonów (często [K + ] na zewnątrz ); 3) otwarte kanały jonowe błony. Zmiany w potencjale błonowym mogą wpływać na konformację białek błonowych, powodując w szczególności otwieranie lub zamykanie kanałów. Jak opisano powyżej, działanie niektórych pomp membranowych zależy od potencjału membranowego. Komórki nerwowe specjalizują się w postrzeganiu zmian potencjału błonowego jako informacji, które muszą być przetwarzane i przekazywane (patrz rozdział 2).
zewnątrzkomórkowe substancje regulujące . Najważniejszym mechanizmem regulacyjnym obejmującym substancje pozakomórkowe jest ich interakcja z określonymi receptorami na błonie komórkowej lub wewnątrz komórki. Substancje te obejmują mediatory synaptyczne, które przekazują informacje między komórkami nerwowymi, czynnikami lokalnymi i substancjami krążącymi we krwi i docierającymi do wszystkich komórek ciała, takimi jak hormony i antygeny. neuroprzekaźniki synaptyczne to małe cząsteczki uwalniane z zakończeń nerwowych w synapsie;
kiedy dotrą do błony plazmatycznej sąsiedniej komórki postsynaptycznej, wyzwalają sygnały elektryczne lub inne mechanizmy regulacyjne. Zagadnienie to zostało szczegółowo omówione w rozdz. 3.
Lokalne środki chemiczne często wydzielane przez wyspecjalizowane komórki. Dyfundują swobodnie w przestrzeni pozakomórkowej, ale ich działanie ogranicza się do niewielkiej grupy komórek ze względu na szybkie niszczenie tych substancji, samoistnie lub pod wpływem enzymów. Jednym z przykładów uwolnienia takich środków jest uwolnienie histamina mastocyty po uszkodzeniu lub odpowiedzi immunologicznej. Histamina powoduje rozluźnienie komórek mięśni gładkich naczyń, zwiększa przepuszczalność śródbłonka naczyniowego i stymuluje zakończenia nerwów czuciowych, które pośredniczą w odczuwaniu swędzenia. Inne lokalne środki chemiczne są wydzielane przez wiele innych komórek. Typowi lokalni agenci to prostaglandyny, stanowiący grupę około 20 pochodnych kwasów tłuszczowych. Są one uwalniane w sposób ciągły z szeroko rozpowszechnionych komórek, ale działają tylko lokalnie, ponieważ są szybko niszczone przez fosfolipazy błonowe. Różne prostaglandyny mają szerokie spektrum działania: mogą wywoływać skurcze komórek mięśni gładkich, powodować agregację płytek krwi lub hamować rozwój ciałka żółtego w jajnikach.
Inni lokalni agenci służą czynniki wzrostowe. Najbardziej znany czynnik wzrostu nerwów (NGF) dla neuronów współczulnych, który jest niezbędny do wzrostu i przeżycia tych neuronów podczas rozwoju in vivo lub w hodowli komórkowej. Oczywiście komórki docelowe dla tej klasy neuronów wydzielają NGF, a tym samym zapewniają prawidłowe unerwienie. Tworząc organy, komórki często muszą „odnajdywać drogę” do komórek docelowych, które mogą znajdować się w znacznych odległościach. W związku z tym musi istnieć wiele wyspecjalizowanych czynników wzrostu, takich jak NGF.
Hormony i antygeny przenoszone przez krew do wszystkich komórek. Antygeny wywołują odpowiedź immunologiczną z komórek niosących określone przeciwciała. Jednak antygeny z reguły są substancjami obcymi, które nie powstają w reagującym organizmie (więcej szczegółów w rozdziale 18). Niektóre hormony, takie jak insulina lub tyroksyna, wpływają na wiele różnych typów komórek, podczas gdy inne, takie jak hormony płciowe, wpływają tylko na niektóre typy komórek. Hormony to albo peptydy, których działanie wyzwalane jest przez ich wiązanie z receptorem na błonie komórkowej, albo steroidy i tyroksyna, które dyfundują przez błonę lipidową i wiążą się z receptorami wewnątrzkomórkowymi. Hormony steroidowe wiążą się z chromatyną jądrową, powodując transkrypcję niektórych genów. Powstałe w wyniku tego białka powodują zmianę funkcji komórkowych, co jest specyficznym działaniem hormonów. Zagadnienia związane z uwalnianiem i działaniem hormonów zostały szczegółowo omówione w rozdz. 17.
Komunikacja wewnątrzkomórkowa z udziałem wtórnych posłańców
Opisane powyżej funkcje regulacyjne obejmują wpływ na błonę komórkową. Informacja otrzymywana przez błonę komórkową musi często powodować reakcję w organellach i jest do nich przenoszona przez różne substancje zwane wtórnymi posłańcami (w przeciwieństwie do pierwszego, docierającego do komórki ze źródeł zewnętrznych). Badanie drugich mediatorów rozwija się szybko i nie ma gwarancji, że obecny poziom zrozumienia problemu będzie wystarczająco pełny. Tutaj dotkniemy trzech dobrze zbadanych mediatorów: Ca 2+ , cAMP i trifosforanu inozytolu.
Wapń.Najprostszym mediatorem wewnątrzkomórkowym jest jon Ca 2+. Jego wolne stężenie w celi spoczynkowej jest bardzo niskie i wynosi 10 -8 -10 -7 mol/l. Może wnikać do komórki przez określone kanały błony, gdy są one otwarte, na przykład, gdy zmienia się potencjał błony (patrz rozdział 2). Wynikający z tego wzrost Ca 2+ wyzwala ważne reakcje w komórce, takie jak skurcz miofibryli, który jest podstawą skurczu mięśni (patrz rozdział 4) lub uwalnianie pęcherzyków zawierających neuroprzekaźniki z zakończeń nerwowych (patrz rozdział 3) . Obie reakcje wymagają stężenia Ca 2+ około 10–5 mol/l. Ca2+, który ma działanie regulacyjne, może być również uwalniany z wewnątrzkomórkowych magazynów, takich jak retikulum endoplazmatyczne. Uwolnienie Ca 2+ z magazynu wymaga udziału innych pośredników (patrz na przykład ryc. 1.16).
Cykliczny monofosforan adenozyny, cAMP. Ostatnio udowodniono, że cykliczny adenozynomonofosforan (cAMP), pochodna głównego źródła energii w organizmie, ATP, jest ważnym drugim przekaźnikiem. Złożony łańcuch reakcji pokazany na ryc. 1.15, zaczyna się od receptora Rs na zewnętrznej powierzchni błony plazmatycznej, która może służyć jako specyficzne miejsce wiązania różnych mediatorów i hormonów. Po związaniu się ze specyficzną „stymulującą” cząsteczką Rs zmienia swoją konformację; te zmiany wpływają na białko Gs na wewnętrznej powierzchni błony w taki sposób, że możliwe staje się jej aktywowanie przez wewnątrzkomórkowy trifosforan guanozyny (GTP). aktywowane białko Gs z kolei stymuluje enzym na wewnętrznej powierzchni błony, cyklazę adenylanową (AC), który katalizuje tworzenie cAMP z ATP. cAMP rozpuszczalny w wodzie i jest mediatorem przenoszącym efekt

Ryż. 1.15.Łańcuch reakcji z udziałem mediatora wewnątrzkomórkowego cAMP (cykliczny monofosforan adenozyny). Zewnętrzne sygnały pobudzające lub hamujące aktywują receptory błonowe R s lub Ri . Receptory te regulują proces wiązania g -białka z wewnątrzkomórkowym GTP (trójfosforanem guanozyny), tym samym stymulujące lub hamujące wewnątrzkomórkową cyklazę adenylanową (AC). Wzmacniający enzym AC przekształca adenozynotrójfosforan (ATP) w cAMP, który jest następnie rozszczepiany do AMP za pomocą fosfodiestera. Wolny cAMP dyfunduje do komórki i aktywuje kinazę adenylanową (kinazę A), uwalniając jej katalityczną podjednostkę C, która katalizuje fosforylację białek wewnątrzkomórkowych, tj. tworzy końcowy efekt bodźca zewnątrzkomórkowego. Schemat pokazuje również leki farmakologiczne i toksyny, które wywołują (+) lub hamują (-) niektóre reakcje (ale ze zmianami)
stymulacja receptora zewnątrzkomórkowego Rs do wewnętrznych struktur komórki.
Równolegle ze stymulującym łańcuchem reakcji obejmujących Rs możliwe wiązanie mediatorów hamujących i hormonów z odpowiednim receptorem R i który ponownie przez białko aktywowane GTP g , hamuje wytwarzanie AC, a tym samym cAMP. Dyfuzując do komórki, cAMP reaguje z kinazą adenylanową (kinazą A); To zwalnia podjednostkę C, która katalizuje fosforylację białek. Ta fosforylacja przekształca białka w ich aktywną formę, a teraz mogą one wywierać swoiste działanie regulacyjne (np. powodować degradację glikogenu). Ten złożony system regulacyjny jest niezwykle wydajny, ponieważ efektem końcowym jest fosforylacja wielu białek, tj. sygnał regulacyjny przechodzi przez łańcuch z dużym współczynnikiem wzmocnienia. Mediatory zewnętrzne, które wiążą się z receptorami R s i R i specyficzne dla każdego z nich są niezwykle różnorodne. Adrenalina, bawiąc się R s lub R i uczestniczy w regulacji metabolizmu lipidów i glikogenu, a także w nasileniu skurczu mięśnia sercowego i innych reakcjach (patrz rozdział 19). Hormon stymulujący tarczycę, aktywacja Rs , stymuluje wydzielanie hormonu tyroksyny przez tarczycę, a prostaglandyna I hamuje agregację płytek krwi. Efekty hamujące, w tym adrenalina, pośredniczone przez R i wyraża się w spowolnieniu lipolizy. W ten sposób, system cAMP jest wielofunkcyjnym wewnątrzkomórkowym systemem regulacyjnym, które mogą być precyzyjnie kontrolowane przez zewnątrzkomórkowe środki stymulujące i hamujące sygnalizację.
Fosforan inozytolu „IF h ”. Wewnątrzkomórkowy układ drugiego mediatora, fosforanu inozytolu, został odkryty dopiero niedawno (ryc. 1.16). W tym przypadku nie ma szlaku hamującego, ale istnieje podobieństwo z układem cAMP, w którym efekt pobudzenia receptora R jest przenoszony na aktywowane GTP białko G na wewnętrznej powierzchni błony. W kolejnym etapie zwykły błonowy fosfatydyloinozytol (PI), który wcześniej otrzymał dwie dodatkowe grupy fosforanowe, jest przekształcany w PI-difosforan (FIF 2), który jest rozszczepiany przez aktywowaną fosfodiesterazę (PDE) do trifosforan inozytolu(IFz) i lipid diacyloglicerol(DAG). Trifosforan inozytolu jest rozpuszczalnym w wodzie drugim przekaźnikiem, który dyfunduje do cytozolu. Działa głównie poprzez uwalnianie Ca 2+ z retikulum endoplazmatycznego. Ca 2+ z kolei działa jako pośrednik, jak opisano powyżej; na przykład aktywuje fosfokinazę zależną od Ca2+, która fosforyluje enzymy. Podjednostka lipidowa DAG (ryc. 1.16) również niesie sygnał, dyfundując w fazie lipidowej błony komórkowej do znajdującej się na jej wewnętrznej powierzchni kinazy C, która jest aktywowana przy udziale fosfatydyloseryny jako kofaktora. Następnie kinaza C wyzwala fosforylację białek, przekształcając je w formę aktywną.
Wewnątrzkomórkowy układ drugiego pośredniczącego interferonu 3 może być również kontrolowany przez różne zewnętrzne mediatory i hormony, w tym acetylocholinę, serotoninę, wazopresynę i hormon stymulujący tarczycę; podobnie jak system cAMP charakteryzuje się różnorodnymi efektami wewnątrzkomórkowymi. Możliwe, że system ten jest również aktywowany przez światło w receptorze wzrokowym oka i odgrywa kluczową rolę w fototransdukcji (patrz rozdział 11). Po raz pierwszy w indywidualnym rozwoju organizmu receptor układu IGF jest aktywowany przez plemniki, w wyniku czego IGF bierze udział w reakcjach regulacyjnych towarzyszących zapłodnieniu komórki jajowej.
Systemy cAMP i IFz-DAG są bardzo skuteczne wzmacniacze biologiczne. Oni są

Ryż. 1.16.Łańcuch reakcji obejmujących wewnątrzkomórkowy interferon pośredni (trójfosforan inozytolu). Podobnie jak w systemie cAMP, w sygnale zewnątrzkomórkowym pośredniczy białkog, który w tym przypadku aktywuje fosfodiesterazę (PDE). Enzym ten rozkłada difosforan fosfatydyloinozyny (FIF) 2 ) w błonie komórkowej przed IF h i diacyloglicerol (DAG); JEŚLI h dyfunduje do cytoplazmy. Tutaj powoduje uwolnienie Ca 2+ z retikulum endoplazmatycznego; wzrost stężenia Ca 2+ w cytoplazmie ([Ca 2+ ] ) aktywuje kinazę białkową, która fosforyluje, a tym samym aktywuje enzymy. Inny produkt, DAG, pozostaje w błonie i aktywuje kinazę białkową C (kofaktor-fosfatydyloseryna, PS). Kinaza białkowa C fosforyluje również enzymy, które pośredniczą w określonych działaniach związanych ze stymulacją receptora zewnętrznego. r . Gałęzie łańcucha reakcji z udziałem IF h i DAG mogą być aktywowane niezależnie odpowiednio przez jonomycynę i ester forbolu (zmodyfikowane)
przekształca reakcję między mediatorem a receptorem błony zewnętrznej w fosforylację wielu białek wewnątrzkomórkowych, co może następnie wpływać na różne funkcje komórki. Jednym z istotnych aspektów problemu jest to, że, o ile dzisiaj wiemy, istnieją tylko te dwa ściśle powiązane ze sobą systemy tego typu, które są wykorzystywane przez licznych mediatorów zewnętrznych do regulacji różnych procesów wewnątrzkomórkowych. Jednocześnie te systemy regulacyjne, w tym Ca 2+ , ściśle ze sobą współdziałają, co pozwala im na precyzyjną regulację funkcji komórkowych.
Poradniki i poradniki
1. Albertowie W., Bray D., Lewis J., Raff M., Roberts DO., Watson J.D.Molecular Biology of the Cell, Nowy Jork i Londyn, Garland Publishing Inc., 1983.
2. Czihak G., Dłuższy H., Ziegler H.(wyd.). Biologia. Berlin, Heidelberg, Nowy Jork, Springer, 1983.
3. hille W. Kanały jonowe błon pobudliwych. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W. Marki H., Ziegler H.(wyd.). Biofizyk. Berlin, Heidelberg, Nowy Jork, Springer, 1984.
5. Jungermanna DO., Mahler H.Biochemia. Berlin, Heidelberg, Nowy Jork, Springer, 1980.
6. Kandel E.R., Schwartz-J.H.,(wyd.). Zasady neuronauki, Nowy Jork, Amsterdam, Oxford, Elsevier, 1985.
7. Schieblera T. H., SchmidtW.Anatomiczny des Menschen. Berlin, Heidelberg, Nowy Jork, Tokio, Springer, 1983.
Oryginalne artykuły i recenzje
8. Berridge M.J. Molekularne podstawy komunikacji w komórce, Sci. Amer 253 124 134 (1985).
9. Berridge M.J., Irvine R.F. Trifosforan inozytolu, nowy drugi przekaźnik w komórkowej transdukcji sygnału. Naturę, 312, 315 321 (1984).
10. Bretscher MS Cząsteczki błony komórkowej, Sci. Amer., 253, 124–134 (1985).
11. Daut J.Żywa komórka jako maszyna przenosząca energię. Minimalny model metabolizmu mięśnia sercowego, Biochem. et Biofizy. Acta, 895, 41-62 (1987).
12. Hodgkin A.L., Katz W. Wpływ jonów sodu na aktywność elektryczną olbrzymiego aksonu kałamarnicy. J Fizjoterapeuta. (Londyn), 108, 37-77 (1949).
13. Hodgkin A.L., Keynes R.D. Aktywny transport kationów w gigantycznych aksonach z Sepia oraz loligo, J Fizjoterapeuta. (Londyn.), 128, 28-42 (1955).
14. Dłuższy P. Kanały jonowe z podstanami konformacyjnymi, Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M. Transport egzoplazmatyczny w układach normalnych i patologicznych. W: Fizjologia i Patologia Aksonów, S.G. Waxman, Wyd. Nowy Jork, Raven Press, 1978.
Zapraszamy do zapoznania się z materiałami oraz.
: błona celulozowa, membrana, cytoplazma z organellami, jądro, wakuole z sokiem komórkowym.Obecność plastydów jest główną cechą komórki roślinnej.
Funkcje ściany komórkowej- determinuje kształt komórki, chroni przed czynnikami środowiskowymi.
błona plazmatyczna- cienka warstwa, składa się z oddziałujących na siebie cząsteczek lipidów i białek, oddziela zawartość wewnętrzną od środowiska zewnętrznego, zapewnia transport wody, substancji mineralnych i organicznych do komórki na drodze osmozy i aktywnego transferu, a także usuwa produkty przemiany materii.
Cytoplazma- wewnętrzne półpłynne środowisko komórki, w którym znajduje się jądro i organelle, zapewnia połączenia między nimi, uczestniczy w głównych procesach życiowych.
Retikulum endoplazmatyczne- sieć rozgałęzionych kanałów w cytoplazmie. Bierze udział w syntezie białek, lipidów i węglowodanów, w transporcie substancji. Rybosomy - ciała znajdujące się na EPS lub w cytoplazmie, składają się z RNA i białka, biorą udział w syntezie białek. EPS i rybosomy to pojedynczy aparat do syntezy i transportu białek.
Mitochondria-organelle oddzielone od cytoplazmy dwiema błonami. Substancje organiczne są w nich utleniane, a cząsteczki ATP są syntetyzowane przy udziale enzymów. Wzrost powierzchni błony wewnętrznej, na której znajdują się enzymy z powodu cristae. ATP to bogata w energię substancja organiczna.
plastydy(chloroplasty, leukoplasty, chromoplasty), ich zawartość w komórce jest główną cechą organizmu roślinnego. Chloroplasty to plastydy zawierające zielony barwnik chlorofil, który pochłania energię świetlną i wykorzystuje ją do syntezy substancji organicznych z dwutlenku węgla i wody. Ograniczenie chloroplastów od cytoplazmy przez dwie błony, liczne wyrostki - grana na błonie wewnętrznej, w której znajdują się cząsteczki chlorofilu i enzymy.
Kompleks Golgiego- system wnęk oddzielonych od cytoplazmy błoną. Nagromadzenie w nich białek, tłuszczów i węglowodanów. Realizacja syntezy tłuszczów i węglowodanów na błonach.
Lizosomy- ciała oddzielone od cytoplazmy pojedynczą błoną. Zawarte w nich enzymy przyspieszają reakcję rozszczepiania złożonych cząsteczek na proste: białka na aminokwasy, węglowodany złożone na proste, lipidy na glicerol i kwasy tłuszczowe, a także niszczą martwe części komórki, całe komórki.
Wakuole- wnęki w cytoplazmie wypełnione sokiem komórkowym, miejsce gromadzenia rezerwowych składników odżywczych, substancji szkodliwych; regulują zawartość wody w komórce.
Rdzeń- główna część komórki, pokryta na zewnątrz podwójną membraną, przebitą porami otoczki jądrowej. Substancje wnikają do rdzenia i są z niego usuwane przez pory. Chromosomy są nośnikami dziedzicznej informacji o cechach organizmu, głównych strukturach jądra, z których każda składa się z jednej cząsteczki DNA w połączeniu z białkami. Jądro jest miejscem syntezy DNA, i-RNA, r-RNA.

Obecność błony zewnętrznej, cytoplazmy z organellami, jądra z chromosomami.
Błona zewnętrzna lub plazmowa- oddziela zawartość komórki od środowiska (inne komórki, substancja międzykomórkowa), składa się z cząsteczek lipidów i białek, zapewnia komunikację między komórkami, transport substancji do wnętrza komórki (pinocytoza, fagocytoza) i poza komórkę.
Cytoplazma- wewnętrzne półpłynne środowisko komórki, które zapewnia komunikację między jądrem a znajdującymi się w nim organellami. Główne procesy życiowej aktywności zachodzą w cytoplazmie.
Organelle komórkowe:
1) retikulum endoplazmatyczne (ER)- układ kanalików rozgałęzionych, zaangażowanych w syntezę białek, lipidów i węglowodanów, w transporcie substancji w komórce;
2) rybosomy- ciała zawierające rRNA znajdują się w ER oraz w cytoplazmie i biorą udział w syntezie białek. EPS i rybosomy to pojedynczy aparat do syntezy i transportu białek;
3) mitochondria- „elektrownie” komórki, oddzielone od cytoplazmy dwiema błonami. Wewnętrzna tworzy cristae (fałdy), które zwiększają jej powierzchnię. Enzymy na cristae przyspieszają reakcje utleniania substancji organicznych i syntezę bogatych w energię cząsteczek ATP;
4) kompleks Golgiego- grupa jam oddzielonych błoną z cytoplazmy, wypełnionych białkami, tłuszczami i węglowodanami, które są albo wykorzystywane w procesach życiowych, albo usuwane z komórki. Błony kompleksu przeprowadzają syntezę tłuszczów i węglowodanów;
5) lizosomy- ciała wypełnione enzymami przyspieszają reakcje rozszczepiania białek na aminokwasy, lipidów na glicerol i kwasów tłuszczowych, polisacharydów na monosacharydy. W lizosomach niszczone są martwe części komórki, całe komórki i komórki.
Wtrącenia komórkowe- Nagromadzenie zapasowych składników odżywczych: białek, tłuszczów i węglowodanów.
Rdzeń- najważniejsza część komórki. Pokryta jest membraną z podwójną błoną z porami, przez które niektóre substancje wnikają do jądra, podczas gdy inne do cytoplazmy. Chromosomy to główne struktury jądra, nośniki dziedzicznej informacji o cechach organizmu. Przenosi się w procesie podziału komórki macierzystej na komórki potomne, a wraz z komórkami zarodkowymi - na organizmy potomne. Jądro jest miejscem syntezy DNA, mRNA, rRNA.
Zadanie:
Wyjaśnij, dlaczego organelle nazywane są wyspecjalizowanymi strukturami komórki?
Odpowiedź: organelle nazywane są wyspecjalizowanymi strukturami komórkowymi, ponieważ pełnią ściśle określone funkcje, informacje dziedziczne są przechowywane w jądrze, ATP jest syntetyzowany w mitochondriach, fotosynteza zachodzi w chloroplastach itp.
Jeśli masz pytania dotyczące cytologii, możesz poprosić o pomoc
Trzecim etapem ewolucji jest pojawienie się komórki.
Cząsteczki białek i kwasów nukleinowych (DNA i RNA) tworzą komórkę biologiczną, najmniejszą jednostkę życia. Komórki biologiczne są „cegiełkami” wszystkich żywych organizmów i zawierają wszystkie materialne kody rozwoju.
Przez długi czas naukowcy uważali budowę komórki za niezwykle prostą. Radziecki słownik encyklopedyczny interpretuje pojęcie komórki w następujący sposób: „Komórka to elementarny system życia, podstawa struktury i życia wszystkich zwierząt i roślin”. Należy zauważyć, że termin „elementarny” w żadnym wypadku nie oznacza „prosty”, wręcz przeciwnie, komórka jest unikalnym fraktalnym tworem Boga, uderzającym swoją złożonością, a jednocześnie wyjątkową spójnością dzieła wszystkich jego elementów.
Kiedy udało nam się zajrzeć do środka za pomocą mikroskopu elektronowego, okazało się, że urządzenie najprostszej komórki jest równie złożone i niezrozumiałe jak sam Wszechświat. Dziś już ustalono, że „Komórka jest szczególną materią Wszechświata, szczególną materią Kosmosu”. Jedna komórka zawiera informacje, które można umieścić w zaledwie kilkudziesięciu tysiącach tomów Wielkiej Encyklopedii Radzieckiej. Tych. komórka to między innymi ogromny „biozbiornik” informacji.
Autor współczesnej teorii ewolucji molekularnej, Manfred Eigen, pisze: „Aby przypadkowo powstała cząsteczka białka, natura musiałaby wykonać około 10130 prób i wydać na to taką liczbę cząsteczek, która wystarczyłaby na 1027 Wszechświaty Jeśli białko zostało zbudowane inteligentnie, to znaczy, że ważność każdego ruchu można było sprawdzić jakimś mechanizmem selekcji, wystarczyło to około 2000 prób. Dochodzimy do paradoksalnego wniosku: program budowy „prymitywnej żywej komórki” jest zakodowany gdzieś na poziomie cząstek elementarnych”.
A jak mogłoby być inaczej. Każda komórka, posiadając DNA, jest obdarzona świadomością, jest świadoma siebie i innych komórek oraz jest w kontakcie ze Wszechświatem, będąc w istocie jego częścią. I chociaż liczba i różnorodność komórek w ludzkim ciele jest niesamowita (około 70 bilionów), wszystkie są do siebie podobne, tak jak wszystkie procesy zachodzące w komórkach są do siebie podobne. Według słów niemieckiego naukowca Rolanda Glasera, konstrukcja komórek biologicznych jest „bardzo przemyślana”. Kto jest dobrze przemyślany?
Odpowiedź jest prosta: białka, kwasy nukleinowe, żywe komórki i wszystkie układy biologiczne są wytworem twórczej aktywności intelektualnego Stwórcy.
Co ciekawe: na poziomie atomowym nie ma różnic między składem chemicznym świata organicznego i nieorganicznego. Innymi słowy, na poziomie atomu komórka powstaje z tych samych elementów, co natura nieożywiona. Różnice znajdują się na poziomie molekularnym. W żywych organizmach oprócz substancji nieorganicznych i wody znajdują się również białka, węglowodany, tłuszcze, kwasy nukleinowe, enzym syntaza ATP i inne niskocząsteczkowe związki organiczne.
Do tej pory komórka została dosłownie rozebrana na atomy w celach badawczych. Nie jest jednak możliwe stworzenie przynajmniej jednej żywej komórki, ponieważ stworzenie komórki oznacza stworzenie cząstki żywego Wszechświata. Akademik W.P. Kaznacheev uważa, że „komórka jest organizmem kosmoplanetarnym… Komórki ludzkie to pewne systemy biozderzaczy eterowo-skrętnych. W tych biozderzaczach zachodzą nieznane nam procesy, zachodzi materializacja kosmicznych form przepływów, ich kosmiczna transformacja cząstki materializują się”.
Woda.
Prawie 80% masy komórek to woda. Według doktora biologii S. Zenina woda, dzięki swojej strukturze klastrowej, jest matrycą informacyjną do zarządzania procesami biochemicznymi. Ponadto to woda jest głównym „celem”, z którym oddziałują oscylacje częstotliwości dźwięku. Uporządkowanie wody komórkowej jest tak wysokie (zbliżone do uporządkowania kryształu), że nazywa się ją ciekłym kryształem.
Wiewiórki.
Białka odgrywają ważną rolę w życiu biologicznym. Komórka zawiera kilka tysięcy białek, które są unikalne dla tego typu komórek (z wyjątkiem komórek macierzystych). Zdolność do syntezy własnych białek jest dziedziczona z komórki na komórkę i utrzymuje się przez całe życie. W trakcie życia komórki białka stopniowo zmieniają swoją strukturę, ich funkcja jest zaburzona. Te zużyte białka są usuwane z komórki i zastępowane nowymi, dzięki czemu zachowana jest żywotna aktywność komórki.
Przede wszystkim zwracamy uwagę na funkcję budulcową białek, ponieważ są one budulcem, z którego składają się błony komórkowe i organelle komórkowe, ściany naczyń krwionośnych, ścięgna, chrząstki itp.
Funkcja sygnalizacyjna białek jest niezwykle interesująca. Okazuje się, że białka mogą pełnić funkcję substancji sygnalizacyjnych, przenoszących sygnały między tkankami, komórkami czy organizmami. Funkcję sygnalizacyjną pełnią białka hormonalne. Komórki mogą komunikować się ze sobą na odległość za pomocą białek sygnałowych przekazywanych przez substancję międzykomórkową.
Białka pełnią również funkcję motoryczną. Wszystkie rodzaje ruchu, do których zdolne są komórki, takie jak skurcze mięśni, są wykonywane przez specjalne białka kurczliwe. Białka pełnią również funkcję transportową. Potrafią przyczepiać różne substancje i przenosić je z jednego miejsca w komórce do drugiego. Na przykład hemoglobina, białko krwi, przyłącza tlen i przenosi go do wszystkich tkanek i narządów ciała. Ponadto białka pełnią również funkcję ochronną. Kiedy obce białka lub komórki są wprowadzane do organizmu, powstają w nim specjalne białka, które wiążą i neutralizują obce komórki i substancje. I wreszcie funkcja energetyczna białek polega na tym, że przy całkowitym rozpadzie 1 g białka uwalniana jest energia w ilości 17,6 kJ.
Struktura komórkowa.
Komórka składa się z trzech nierozerwalnie połączonych części: błony, cytoplazmy i jądra, a budowa i funkcja jądra w różnych okresach życia komórki są różne. Na życie komórki składają się dwa okresy: podział, w wyniku którego powstają dwie komórki potomne, oraz okres między podziałami, zwany interfazą.
Błona komórkowa bezpośrednio oddziałuje ze środowiskiem zewnętrznym i oddziałuje z sąsiednimi komórkami. Składa się z warstwy zewnętrznej i znajdującej się pod nią błony plazmatycznej. Warstwa powierzchniowa komórek zwierzęcych nazywana jest glikokaliami. Łączy komórki ze środowiskiem zewnętrznym i wszystkimi otaczającymi je substancjami. Jego grubość to mniej niż 1 mikron.
Struktura komórkowa
Błona komórkowa jest bardzo ważną częścią komórki. Łączy wszystkie składniki komórkowe i ogranicza środowisko zewnętrzne i wewnętrzne.
Między komórkami a środowiskiem zewnętrznym zachodzi ciągła wymiana substancji. Woda, różne sole w postaci pojedynczych jonów, cząsteczki nieorganiczne i organiczne dostają się do komórki ze środowiska zewnętrznego. Produkty metabolizmu, a także substancje syntetyzowane w komórce: białka, węglowodany, hormony, które są wytwarzane w komórkach różnych gruczołów, są wydalane do środowiska zewnętrznego przez błonę komórki. Transport substancji jest jedną z głównych funkcji błony plazmatycznej.
Cytoplazma- wewnętrzne półpłynne medium, w którym zachodzą główne procesy metaboliczne. Ostatnie badania wykazały, że cytoplazma nie jest rodzajem roztworu, którego składniki oddziałują ze sobą w przypadkowych zderzeniach. Można go porównać do galaretki, która zaczyna „drżeć” w odpowiedzi na wpływy zewnętrzne. W ten sposób cytoplazma odbiera i przekazuje informacje.
Jądro i różne organelle znajdują się w cytoplazmie, które są przez nią połączone w jedną całość, co zapewnia ich interakcję i aktywność komórki jako jednego integralnego układu. Jądro znajduje się w centralnej części cytoplazmy. Cała wewnętrzna strefa cytoplazmy jest wypełniona siateczką endoplazmatyczną, która jest organoidem komórkowym: układem kanalików, pęcherzyków i „cystern” ograniczonych błonami. Retikulum endoplazmatyczne bierze udział w procesach metabolicznych, zapewniając transport substancji ze środowiska do cytoplazmy oraz pomiędzy poszczególnymi strukturami wewnątrzkomórkowymi, ale jego główną funkcją jest udział w syntezie białek, która odbywa się w rybosomach. - mikroskopijne ciałka o okrągłym kształcie o średnicy 15-20 nm. Zsyntetyzowane białka są najpierw gromadzone w kanałach i jamach retikulum endoplazmatycznego, a następnie transportowane do organelli i miejsc komórkowych, gdzie są konsumowane.
Oprócz białek cytoplazma zawiera również mitochondria, małe ciała o wielkości 0,2-7 mikrona, które nazywane są „elektrowniami” komórek. Reakcje redoks zachodzą w mitochondriach, dostarczając komórkom energii. Liczba mitochondriów w jednej komórce waha się od kilku do kilku tysięcy.
Rdzeń- żywotna część komórki, kontroluje syntezę białek, a za ich pośrednictwem wszystkie procesy fizjologiczne w komórce. W jądrze niedzielącej się komórki rozróżnia się błonę jądrową, sok jądrowy, jąderko i chromosomy. Poprzez otoczkę jądrową zachodzi ciągła wymiana substancji między jądrem a cytoplazmą. Pod otoczką jądrową - sok jądrowy (substancja półpłynna), który zawiera jąderko i chromosomy. Jąderko jest gęstym, zaokrąglonym ciałem, którego wymiary mogą się znacznie różnić, od 1 do 10 mikronów i więcej. Składa się głównie z rybonukleoprotein; uczestniczy w tworzeniu rybosomów. Zwykle w komórce znajduje się 1-3 jąderek, czasem nawet kilkaset. Jąderko składa się z RNA i białka.
Wraz z nadejściem komórki na Ziemi powstało Życie!
Ciąg dalszy nastąpi...


