Menyediakan hubungan sel dengan lingkungan. Hubungan organisme dengan lingkungan
Hubungan organisme dengan lingkungan, dari sudut pandang fisikokimia, adalah sistem terbuka, yaitu sistem di mana proses biokimia sedang berlangsung. Zat-zat awal berasal dari lingkungan, dan zat-zat yang juga terus menerus terbentuk dikeluarkan. Keseimbangan antara laju dan konsentrasi produk reaksi multiarah dalam tubuh adalah kondisional, imajiner, karena asupan dan pengeluaran zat tidak berhenti. Koneksi terus menerus dengan lingkungan dan memungkinkan kita untuk menganggap organisme hidup sebagai sistem terbuka.
Matahari adalah sumber energi bagi semua sel hidup. Sel tumbuhan menangkap energi sinar matahari dengan bantuan klorofil, menggunakannya untuk reaksi asimilasi selama fotosintesis. Sel hewan, jamur, bakteri menggunakan energi matahari secara tidak langsung, ketika membelah zat organik yang disintesis oleh tanaman duniawi.
Bagian dari nutrisi sel dipecah dalam proses respirasi sel, sehingga memasok energi yang diperlukan untuk berbagai jenis aktivitas seluler. Proses ini terjadi di organel yang disebut mitokondria. Mitokondria terdiri dari dua membran: membran luar, yang memisahkan organel dari sitoplasma, dan membran dalam, yang membentuk banyak lipatan. Produk utama respirasi adalah ATP. Ia meninggalkan mitokondria dan digunakan sebagai sumber energi untuk banyak reaksi kimia di sitoplasma dan membran sel. Jika oksigen diperlukan untuk pelaksanaan respirasi sel, maka respirasi disebut respirasi aerob, tetapi jika reaksi berlangsung tanpa oksigen, maka kita berbicara tentang respirasi anaerob.
Untuk setiap jenis pekerjaan yang dilakukan dalam sel, energi digunakan dalam satu bentuk - dalam bentuk energi dari ikatan fosfat ATP. ATP adalah senyawa yang sangat mobile. Pembentukan ATP terjadi di membran dalam mitokondria. ATP disintesis di semua sel selama respirasi karena energi oksidasi karbohidrat, lemak dan zat organik lainnya. Dalam sel tumbuhan hijau, jumlah utama ATP disintesis dalam kloroplas karena energi matahari. Di dalamnya, selama fotosintesis, lebih banyak ATP diproduksi daripada di mitokondria. ATP terurai dengan pemutusan ikatan fosfor-oksigen dan pelepasan energi. Ini terjadi di bawah aksi enzim ATPase dalam proses hidrolisis ATP - penambahan air dengan penghapusan molekul asam fosfat. Akibatnya, ATP diubah menjadi ADP, dan jika dua molekul asam fosfat dipecah, maka menjadi AMP. Reaksi pembelahan setiap gram molekul asam disertai dengan pelepasan 40 kJ. Ini adalah hasil energi yang sangat besar, sehingga ikatan fosfor-oksigen ATP biasa disebut makroergik (energi tinggi).
Penggunaan ATP dalam reaksi pertukaran plastik dilakukan dengan konjugasinya dengan hidrolisis ATP. Molekul berbagai zat diisi dengan energi dengan mengikat gugus fosfor yang dilepaskan selama hidrolisis dari molekul ATP, yaitu melalui fosforilasi.
Ciri turunan fosfat adalah bahwa mereka tidak dapat meninggalkan sel, meskipun bentuk "terlepas" mereka bebas melewati membran. Karena itu, molekul terfosforilasi tetap berada di dalam sel sampai mereka digunakan dalam reaksi yang sesuai.
Proses kebalikan dari mengubah ADP menjadi ATP terjadi dengan menempelkan molekul asam fosfat ke ADP, melepaskan air dan menyerap sejumlah besar energi.
Dengan demikian, ATP adalah sumber energi universal dan langsung untuk aktivitas sel. Ini menciptakan dana energi seluler tunggal dan memungkinkan untuk mendistribusikan dan mengangkutnya dari satu bagian sel ke bagian lain.
Transfer gugus fosfat memainkan peran penting dalam reaksi kimia seperti perakitan makromolekul dari monomer. Misalnya, asam amino hanya dapat digabungkan menjadi peptida jika sebelumnya telah difosforilasi. Proses mekanis dari kontraksi atau gerakan, transfer zat terlarut melawan gradien konsentrasi, dan proses lainnya terkait dengan pengeluaran energi yang tersimpan dalam ATP.
Proses pertukaran energi dapat direpresentasikan sebagai berikut. Zat organik bermolekul tinggi dalam sitoplasma secara enzimatik, dengan hidrolisis, diubah menjadi yang lebih sederhana, yang terdiri dari: protein - menjadi asam amino, poli- dan disakarida - menjadi monosakarida (+ glukosa), lemak menjadi gliserol dan asam lemak. Proses oksidatif tidak ada, sedikit energi yang dilepaskan, yang tidak digunakan dan masuk ke bentuk termal. Sebagian besar sel menggunakan karbohidrat terlebih dahulu. Polisakarida (pati pada tumbuhan dan glikogen pada hewan) dihidrolisis menjadi glukosa. Oksidasi glukosa terjadi dalam tiga fase: glikolisis, dekarboksilasi oksidatif (siklus Krebs - siklus asam sitrat) dan fosforilasi oksidatif (rantai pernapasan). Glikolisis, sebagai akibatnya satu molekul glukosa dipecah menjadi dua molekul asam piruvat dengan pelepasan dua molekul ATP, terjadi di sitoplasma. Dengan tidak adanya oksigen, asam piruvat diubah menjadi etanol (fermentasi) atau asam laktat (respirasi anaerob).
Jika glikolisis dilakukan dalam sel hewan, molekul glukosa enam karbon terurai menjadi dua molekul asam laktat. Proses ini multi-tahap. Ini dilakukan secara berurutan oleh 13 enzim. Selama fermentasi alkohol, dua molekul etanol dan dua molekul CO2 terbentuk dari molekul glukosa.
Glikolisis adalah fase umum untuk respirasi anaerobik dan aerobik, dua lainnya dilakukan hanya dalam kondisi aerobik. Proses oksidasi bebas oksigen, di mana hanya sebagian energi metabolit yang dilepaskan dan digunakan, merupakan proses akhir bagi organisme anaerob. Dengan adanya oksigen, asam piruvat masuk ke mitokondria, di mana, sebagai hasil dari sejumlah reaksi yang berurutan, asam piruvat dioksidasi sempurna secara aerobik menjadi H2O dan CO2 dengan fosforilasi ADP menjadi ATP secara simultan. Pada saat yang sama, glikolisis memberikan dua molekul ATP, dua - siklus Krebs, 34 - rantai pernapasan. Hasil bersih dari oksidasi lengkap satu molekul glukosa menjadi H2O dan CO2 adalah 38 molekul.
Jadi, dalam organisme aerobik, penguraian akhir zat organik dilakukan dengan mengoksidasinya dengan oksigen atmosfer menjadi zat anorganik sederhana: CO2 dan H2O. Proses ini terjadi pada krista mitokondria. Dalam hal ini, jumlah maksimum energi bebas dilepaskan, sebagian besar disimpan dalam molekul ATP. Sangat mudah untuk melihat bahwa oksidasi aerobik menyediakan sel dengan energi bebas secara maksimal.
Sebagai hasil dari katabolisme, molekul ATP yang kaya energi menumpuk di dalam sel, dan CO2 dan kelebihan air dilepaskan ke lingkungan eksternal.
Molekul gula yang tidak diperlukan untuk respirasi dapat disimpan di dalam sel. Kelebihan lipid akan dibelah, setelah itu produk pembelahannya masuk ke mitokondria sebagai substrat untuk respirasi, atau disimpan sebagai cadangan di sitoplasma dalam bentuk tetesan lemak. Protein dibangun dari asam amino yang masuk ke dalam sel. Sintesis protein terjadi di organel yang disebut ribosom. Setiap ribosom terdiri dari dua subpartikel - besar dan kecil: kedua subpartikel termasuk molekul protein dan molekul RNA.
Ribosom sering melekat pada sistem membran khusus, yang terdiri dari tangki dan vesikel, pada apa yang disebut retikulum endoplasma (ER); dalam sel yang menghasilkan banyak protein, retikulum endoplasma seringkali berkembang sangat baik dan penuh dengan ribosom. Beberapa enzim hanya efektif jika mereka melekat pada membran. Sebagian besar enzim yang terlibat dalam sintesis lipid terletak di sini. Jadi, retikulum endoplasma, seolah-olah, semacam desktop sel.
Selain itu, RE membagi sitoplasma menjadi bagian-bagian yang terpisah, atau kompartemen, yaitu, memisahkan berbagai proses kimia yang terjadi secara bersamaan di sitoplasma, dan dengan demikian mengurangi kemungkinan bahwa proses-proses ini akan saling mengganggu.
Seringkali produk yang dibentuk oleh sel tertentu digunakan di luar sel. Dalam kasus seperti itu, protein yang disintesis pada ribosom melewati membran retikulum endoplasma dan dikemas ke dalam vesikel membran yang terbentuk di sekitar mereka, yang kemudian diikat dari RE. Gelembung ini, merata dan menumpuk satu sama lain, seperti pancake dalam tumpukan, membentuk struktur karakteristik yang disebut kompleks Golgi, atau aparatus Golgi. Selama mereka tinggal di aparatus Golgi, protein mengalami perubahan tertentu. Ketika saatnya bagi mereka untuk meninggalkan sel, vesikel membran bergabung dengan membran sel dan mengosongkan, menuangkan isinya ke luar, yaitu sekresi terjadi dengan eksositosis.
Lisosom juga terbentuk di aparatus Golgi - kantung membran yang mengandung enzim pencernaan. Memahami bagaimana sel membuat, mengemas, dan mengekspor protein tertentu, dan bagaimana sel "mengetahui" protein mana yang harus disimpan untuk dirinya sendiri, adalah salah satu cabang sitologi modern yang paling menarik.
Selaput sel apa pun terus bergerak dan berubah. Membran ER bergerak perlahan di seluruh sel. Bagian terpisah dari membran ini dipisahkan dan membentuk vesikel, yang sementara menjadi bagian dari aparatus Golgi, dan kemudian, dalam proses eksositosis, bergabung dengan membran sel.
Kemudian, bahan membran kembali ke sitoplasma, di mana ia digunakan kembali.
Pertukaran zat yang masuk ke dalam sel atau yang dikeluarkan olehnya ke luar, serta pertukaran berbagai sinyal dengan lingkungan mikro dan makro, terjadi melalui membran luar sel. Seperti diketahui, membran sel adalah lapisan ganda lipid di mana berbagai molekul protein tertanam yang bertindak sebagai reseptor khusus, saluran ion, perangkat yang secara aktif mentransfer atau menghilangkan berbagai bahan kimia, kontak antar sel, dll. Dalam sel eukariotik yang sehat, fosfolipid didistribusikan di membran asimetris: permukaan luar terdiri dari sphingomyelin dan phosphatidylcholine, permukaan bagian dalam terdiri dari phosphatidylserine dan phosphatidylethanolamine. Mempertahankan asimetri seperti itu membutuhkan pengeluaran energi. Oleh karena itu, dalam kasus kerusakan sel, infeksinya, kekurangan energi, permukaan luar membran diperkaya dengan fosfolipid yang tidak biasa, yang menjadi sinyal bagi sel dan enzim lain tentang kerusakan sel dengan reaksi yang sesuai terhadapnya. Peran paling penting dimainkan oleh bentuk larut fosfolipase A2, yang memecah asam arakidonat dan menciptakan lisoform dari fosfolipid di atas. Asam arakidonat adalah link pembatas untuk penciptaan mediator inflamasi seperti eikosanoid, dan molekul pelindung - pentraxins (C-reactive protein (CRP), prekursor protein amiloid) - melekat pada lisoform di membran, diikuti oleh aktivasi melengkapi sistem sepanjang jalur klasik dan penghancuran sel.
Struktur membran berkontribusi pada pelestarian fitur lingkungan internal sel, perbedaannya dari lingkungan eksternal. Ini dipastikan oleh permeabilitas selektif membran sel, adanya mekanisme transpor aktif di dalamnya. Pelanggaran mereka sebagai akibat dari kerusakan langsung, misalnya, oleh tetrodotoxin, ouabain, tetraethylammonium, atau dalam kasus pasokan energi yang tidak mencukupi dari "pompa" yang sesuai, menyebabkan pelanggaran komposisi elektrolit sel, perubahan metabolismenya. , pelanggaran fungsi spesifik - kontraksi, konduksi impuls eksitasi, dll. Pelanggaran saluran ion seluler (kalsium, natrium, kalium, dan klorida) pada manusia juga dapat ditentukan secara genetik oleh mutasi gen yang bertanggung jawab atas struktur ini saluran. Yang disebut channelopathies adalah penyebab penyakit keturunan pada sistem saraf, otot, dan pencernaan. Asupan air yang berlebihan di dalam sel dapat menyebabkan pecahnya - sitolisis - karena perforasi membran selama aktivasi komplemen atau serangan oleh limfosit sitotoksik dan pembunuh alami.
Banyak reseptor dibangun ke dalam membran sel - struktur yang, bila digabungkan dengan molekul sinyal spesifik yang sesuai (ligan), mengirimkan sinyal ke dalam sel. Ini terjadi melalui berbagai kaskade regulasi, yang terdiri dari molekul aktif enzimatik, yang diaktifkan secara berurutan dan pada akhirnya berkontribusi pada implementasi berbagai program seluler, seperti pertumbuhan dan proliferasi, diferensiasi, motilitas, penuaan, dan kematian sel. Kaskade regulasi cukup banyak, tetapi jumlahnya belum sepenuhnya ditentukan. Sistem reseptor dan kaskade regulasi yang terkait dengannya juga ada di dalam sel; mereka membuat jaringan pengaturan tertentu dengan titik konsentrasi, distribusi, dan pilihan jalur sinyal lebih lanjut tergantung pada keadaan fungsional sel, tahap perkembangannya, dan aksi simultan sinyal dari reseptor lain. Hasil dari ini mungkin penghambatan atau penguatan sinyal, arahnya di sepanjang jalur regulasi yang berbeda. Baik aparatus reseptor maupun jalur transduksi sinyal melalui kaskade regulasi, misalnya ke nukleus, dapat terganggu sebagai akibat dari defek genetik yang terjadi sebagai defek kongenital pada tingkat organisme atau karena mutasi somatik pada sel tertentu. Tipe. Mekanisme ini dapat dirusak oleh agen infeksius, toksin, dan juga berubah selama penuaan. Tahap terakhir ini mungkin merupakan pelanggaran fungsi sel, proses proliferasi dan diferensiasinya.
Molekul yang berperan penting dalam proses interaksi antar sel juga terdapat pada permukaan sel. Ini mungkin termasuk protein adhesi sel, antigen kompatibilitas jaringan, spesifik jaringan, antigen pembeda, dll. Perubahan komposisi molekul ini menyebabkan pelanggaran interaksi antar sel dan dapat menyebabkan aktivasi mekanisme yang sesuai untuk eliminasi sel tersebut, karena mereka menimbulkan bahaya tertentu bagi integritas tubuh sebagai reservoir infeksi, terutama virus, atau sebagai inisiator potensial pertumbuhan tumor.
Pelanggaran pasokan energi sel
Sumber energi dalam sel adalah makanan, setelah pemecahan energi yang dilepaskan ke zat akhir. Mitokondria adalah tempat utama produksi energi, di mana zat dioksidasi dengan bantuan enzim rantai pernapasan. Oksidasi adalah pemasok energi utama, karena sebagai hasil glikolisis, tidak lebih dari 5% energi dilepaskan dari jumlah substrat oksidasi (glukosa) yang sama, dibandingkan dengan oksidasi. Sekitar 60% energi yang dilepaskan selama oksidasi diakumulasikan oleh fosforilasi oksidatif dalam fosfat makroergik (ATP, kreatin fosfat), sisanya dikeluarkan sebagai panas. Di masa depan, fosfat berenergi tinggi digunakan oleh sel untuk proses seperti pemompaan, sintesis, pembelahan, pergerakan, sekresi, dll. Ada tiga mekanisme, yang kerusakannya dapat menyebabkan gangguan pasokan energi ke sel : yang pertama adalah mekanisme sintesis enzim metabolisme energi, yang kedua adalah mekanisme fosforilasi oksidatif, yang ketiga - mekanisme penggunaan energi.
Pelanggaran transpor elektron dalam rantai pernapasan mitokondria atau pelepasan oksidasi dan fosforilasi ADP dengan hilangnya potensi proton - kekuatan pendorong generasi ATP, menyebabkan melemahnya fosforilasi oksidatif sedemikian rupa sehingga sebagian besar energi dihamburkan dalam bentuk panas dan jumlah senyawa makroergik berkurang. Pemisahan oksidasi dan fosforilasi di bawah pengaruh adrenalin digunakan oleh sel-sel organisme homoiotermik untuk meningkatkan produksi panas sambil mempertahankan suhu tubuh yang konstan selama pendinginan atau peningkatannya selama demam. Perubahan signifikan dalam struktur mitokondria dan metabolisme energi diamati pada tirotoksikosis. Perubahan ini awalnya reversibel, tetapi setelah titik tertentu mereka menjadi ireversibel: mitokondria fragmen, hancur atau membengkak, kehilangan krista, berubah menjadi vakuola, dan akhirnya menumpuk zat seperti hialin, feritin, kalsium, lipofuscin. Pada pasien dengan penyakit kudis, mitokondria menyatu untuk membentuk kondrosfer, kemungkinan karena kerusakan membran oleh senyawa peroksida. Kerusakan signifikan pada mitokondria terjadi di bawah pengaruh radiasi pengion, selama transformasi sel normal menjadi sel ganas.
Mitokondria adalah gudang ion kalsium yang kuat, di mana konsentrasinya beberapa kali lipat lebih tinggi daripada di sitoplasma. Ketika mitokondria rusak, kalsium memasuki sitoplasma, menyebabkan aktivasi proteinase dengan kerusakan struktur intraseluler dan gangguan fungsi sel yang sesuai, misalnya kontraktur kalsium atau bahkan "kematian kalsium" di neuron. Sebagai akibat dari pelanggaran kemampuan fungsional mitokondria, pembentukan senyawa peroksida radikal bebas meningkat tajam, yang memiliki reaktivitas sangat tinggi dan karenanya merusak komponen sel penting - asam nukleat, protein, dan lipid. Fenomena ini diamati selama apa yang disebut stres oksidatif dan dapat memiliki konsekuensi negatif bagi keberadaan sel. Dengan demikian, kerusakan pada membran luar mitokondria disertai dengan pelepasan ke dalam sitoplasma zat yang terkandung dalam ruang antarmembran, terutama sitokrom C dan beberapa zat aktif biologis lainnya, yang memicu reaksi berantai yang menyebabkan kematian sel terprogram - apoptosis. Dengan merusak DNA mitokondria, reaksi radikal bebas mendistorsi informasi genetik yang diperlukan untuk pembentukan enzim rantai pernapasan tertentu yang diproduksi secara khusus di mitokondria. Hal ini menyebabkan gangguan yang lebih besar dari proses oksidatif. Secara keseluruhan, perangkat genetik intrinsik mitokondria, dibandingkan dengan perangkat genetik nukleus, kurang terlindungi dari pengaruh berbahaya yang mampu mengubah informasi genetik yang dikodekan di dalamnya. Akibatnya, disfungsi mitokondria terjadi sepanjang hidup, misalnya, dalam proses penuaan, selama transformasi ganas sel, serta dengan latar belakang penyakit mitokondria herediter yang terkait dengan mutasi DNA mitokondria dalam telur. Saat ini, lebih dari 50 mutasi mitokondria telah dijelaskan yang menyebabkan penyakit degeneratif herediter pada sistem saraf dan otot. Mereka ditransmisikan ke anak secara eksklusif dari ibu, karena mitokondria sperma bukan bagian dari zigot dan, karenanya, organisme baru.
Pelanggaran pelestarian dan transmisi informasi genetik
Inti sel mengandung sebagian besar informasi genetik dan dengan demikian memastikan fungsi normalnya. Dengan bantuan ekspresi gen selektif, ia mengoordinasikan kerja sel dalam interfase, menyimpan informasi genetik, menciptakan kembali, dan mentransfer materi genetik dalam proses pembelahan sel. Replikasi DNA dan transkripsi RNA terjadi di dalam nukleus. Berbagai faktor patogen, seperti ultraviolet dan radiasi pengion, oksidasi radikal bebas, bahan kimia, virus, dapat merusak DNA. Diperkirakan setiap sel hewan berdarah panas dalam 1 hari. kehilangan lebih dari 10.000 basis. Untuk ini harus ditambahkan pelanggaran saat menyalin selama pembagian. Jika kerusakan ini berlanjut, sel tidak akan mampu bertahan. Perlindungan terletak pada keberadaan sistem perbaikan yang kuat, seperti endonuklease ultraviolet, replikasi reparatif dan sistem perbaikan rekombinasi, yang menggantikan kerusakan DNA. Cacat genetik dalam sistem reparatif menyebabkan perkembangan penyakit karena peningkatan kepekaan terhadap faktor perusak DNA. Ini adalah xeroderma berpigmen, serta beberapa sindrom penuaan yang dipercepat, disertai dengan peningkatan kecenderungan terjadinya tumor ganas.
Sistem pengaturan proses replikasi DNA, transkripsi messenger RNA (mRNA), translasi informasi genetik dari asam nukleat ke dalam struktur protein cukup kompleks dan bertingkat. Selain kaskade regulasi yang memicu aksi lebih dari 3000 faktor transkripsi yang mengaktifkan gen tertentu, ada juga sistem regulasi bertingkat yang dimediasi oleh molekul RNA kecil (RNA pengganggu; RNAi). Genom manusia, yang terdiri dari sekitar 3 miliar basa purin dan pirimidin, hanya mengandung 2% gen struktural yang bertanggung jawab untuk sintesis protein. Sisanya menyediakan sintesis RNA pengatur, yang, bersama-sama dengan faktor transkripsi, mengaktifkan atau memblokir kerja gen struktural pada tingkat DNA dalam kromosom atau mempengaruhi translasi messenger RNA (mRNA) selama pembentukan molekul polipeptida di sitoplasma. . Pelanggaran informasi genetik dapat terjadi baik pada tingkat gen struktural dan bagian pengatur DNA dengan manifestasi yang sesuai dalam bentuk berbagai penyakit keturunan.
Baru-baru ini, banyak perhatian telah diberikan pada perubahan materi genetik yang terjadi selama perkembangan individu suatu organisme dan terkait dengan penghambatan atau aktivasi bagian DNA dan kromosom tertentu karena metilasi, asetilasi, dan fosforilasinya. Perubahan-perubahan ini berlangsung lama, kadang-kadang sepanjang hidup organisme dari embriogenesis hingga usia tua, dan disebut pewarisan epigenomik.
Reproduksi sel dengan informasi genetik yang diubah juga dicegah oleh sistem (faktor) kontrol siklus mitosis. Mereka berinteraksi dengan cyclin-dependent protein kinase dan subunit katalitiknya - cyclin - dan memblokir perjalanan siklus mitosis lengkap oleh sel, menghentikan pembelahan di perbatasan antara fase prasintetis dan sintetik (blok G1 / S) sampai perbaikan DNA selesai , dan jika tidak mungkin, mereka memulai sel kematian terprogram. Faktor-faktor ini termasuk gen p53, yang mutasinya menyebabkan hilangnya kendali atas proliferasi sel yang diubah; itu terjadi pada hampir 50% kanker manusia. Pos pemeriksaan kedua dari perjalanan siklus mitosis terletak di perbatasan G2/M. Di sini, distribusi materi kromosom yang benar antara sel anak dalam mitosis atau meiosis dikendalikan menggunakan mekanisme kompleks yang mengontrol gelendong sel, pusat dan sentromer (kinetokor). Ketidakefisienan mekanisme ini menyebabkan pelanggaran distribusi kromosom atau bagian-bagiannya, yang dimanifestasikan oleh tidak adanya kromosom di salah satu sel anak (aneuploidi), adanya kromosom ekstra (poliploidi), pemisahan bagian dari kromosom (penghapusan) dan transfernya ke kromosom lain (translokasi). Proses seperti itu sangat sering diamati selama reproduksi sel yang mengalami degenerasi dan transformasi ganas. Jika ini terjadi selama meiosis dengan sel germinal, itu mengarah pada kematian janin pada tahap awal perkembangan embrio, atau kelahiran organisme dengan penyakit kromosom.
Reproduksi sel yang tidak terkontrol selama pertumbuhan tumor terjadi sebagai akibat dari mutasi pada gen yang mengontrol proliferasi sel dan disebut onkogen. Di antara lebih dari 70 onkogen yang dikenal saat ini, sebagian besar merupakan komponen pengatur pertumbuhan sel, beberapa merupakan faktor transkripsi yang mengatur aktivitas gen, serta faktor penghambat pembelahan dan pertumbuhan sel. Faktor lain yang membatasi ekspansi (penyebaran) berlebihan dari sel yang berproliferasi adalah pemendekan ujung kromosom - telomer, yang tidak dapat bereplikasi sepenuhnya sebagai akibat dari interaksi sterik murni, oleh karena itu, setelah setiap pembelahan sel, telomer dipersingkat oleh bagian tertentu dari pangkalan. Jadi, sel-sel yang berkembang biak dari organisme dewasa, setelah sejumlah pembelahan tertentu (biasanya dari 20 hingga 100, tergantung pada jenis organisme dan usianya), menghabiskan panjang telomer dan replikasi kromosom selanjutnya berhenti. Fenomena ini tidak terjadi pada epitel spermatogenik, enterosit dan sel embrio karena adanya enzim telomerase, yang mengembalikan panjang telomer setelah setiap pembelahan. Di sebagian besar sel organisme dewasa, telomerase diblokir, tetapi sayangnya, telomerase diaktifkan di sel tumor.
Hubungan antara nukleus dan sitoplasma, pengangkutan zat di kedua arah dilakukan melalui pori-pori di membran nuklir dengan partisipasi sistem transportasi khusus dengan konsumsi energi. Dengan demikian, energi dan zat plastik, molekul sinyal (faktor transkripsi) diangkut ke nukleus. Aliran balik membawa molekul mRNA ke dalam sitoplasma dan mentransfer RNA (tRNA), ribosom yang diperlukan untuk sintesis protein dalam sel. Cara pengangkutan zat yang sama melekat pada virus, khususnya, seperti HIV. Mereka mentransfer materi genetik mereka ke nukleus sel inang dengan inklusi lebih lanjut dalam genom inang dan transfer RNA virus yang baru terbentuk ke dalam sitoplasma untuk sintesis protein lebih lanjut dari partikel virus baru.
Pelanggaran proses sintesis
Proses sintesis protein terjadi di tangki retikulum endoplasma, berhubungan erat dengan pori-pori di membran nuklir, melalui mana ribosom, tRNA dan mRNA memasuki retikulum endoplasma. Di sini, sintesis rantai polipeptida dilakukan, yang kemudian memperoleh bentuk akhirnya di retikulum endoplasma agranular dan kompleks pipih (kompleks Golgi), di mana mereka menjalani modifikasi pasca-translasi dan asosiasi dengan molekul karbohidrat dan lipid. Molekul protein yang baru terbentuk tidak tetap berada di tempat sintesis, tetapi dengan bantuan proses pengaturan yang kompleks, yang disebut protein kinesis, secara aktif ditransfer ke bagian sel yang terisolasi di mana mereka akan melakukan fungsi yang dimaksudkan. Dalam hal ini, langkah yang sangat penting adalah penataan molekul yang ditransfer ke dalam konfigurasi spasial yang sesuai yang mampu melakukan fungsi bawaannya. Penataan seperti itu terjadi dengan bantuan enzim khusus atau pada matriks molekul protein khusus - pendamping, yang membantu molekul protein, yang baru terbentuk atau diubah karena pengaruh eksternal, memperoleh struktur tiga dimensi yang benar. Dalam kasus efek buruk pada sel, bila ada kemungkinan pelanggaran struktur molekul protein (misalnya, dengan peningkatan suhu tubuh, proses infeksi, keracunan), konsentrasi pendamping dalam sel meningkat tajam. Oleh karena itu, molekul seperti itu juga disebut protein stres, atau protein kejutan panas. Pelanggaran struktur molekul protein mengarah pada pembentukan konglomerat inert kimia yang disimpan di dalam sel atau di luarnya dalam kasus amiloidosis, penyakit Alzheimer, dll. Kadang-kadang molekul analog pra-terstruktur dapat berfungsi sebagai matriks, dan dalam dalam hal ini, jika penataan primer tidak terjadi dengan benar, semua molekul berikutnya juga akan rusak. Situasi ini terjadi pada apa yang disebut penyakit prion (scrappie pada domba, rabies pada sapi, kuru, penyakit Creutzfeldt-Jakob pada manusia), ketika cacat pada salah satu protein membran sel saraf menyebabkan akumulasi massa inert berikutnya di dalam. sel dan gangguan aktivitas vitalnya.
Pelanggaran proses sintesis dalam sel dapat terjadi pada berbagai tahap: transkripsi RNA dalam nukleus, translasi polipeptida dalam ribosom, modifikasi pasca-translasi, hipermetilasi dan glikosilasi molekul beige, transportasi dan distribusi protein dalam sel dan penghapusannya. ke luar. Dalam hal ini, seseorang dapat mengamati peningkatan atau penurunan jumlah ribosom, pemecahan poliribosom, perluasan tangki retikulum endoplasma granular, hilangnya ribosom olehnya, pembentukan vesikel dan vakuola. Jadi, dalam kasus keracunan jamur payung pucat, enzim RNA polimerase rusak, yang mengganggu transkripsi. Toksin difteri, menonaktifkan faktor pemanjangan, mengganggu proses translasi, menyebabkan kerusakan pada miokardium. Alasan pelanggaran sintesis beberapa molekul protein tertentu dapat menjadi agen infeksi. Sebagai contoh, virus herpes menghambat sintesis dan ekspresi molekul antigen MHC, yang memungkinkan mereka untuk sebagian menghindari kontrol kekebalan, dan basil wabah menghambat sintesis mediator peradangan akut. Munculnya protein yang tidak biasa dapat menghentikan pemecahan lebih lanjut dan menyebabkan akumulasi bahan lembam atau bahkan beracun. Sampai batas tertentu, gangguan proses pembusukan juga dapat berkontribusi pada hal ini.
Pelanggaran proses pembusukan
Bersamaan dengan sintesis protein di dalam sel, pembusukannya terus terjadi. Dalam kondisi normal, ini memiliki signifikansi pengaturan dan formatif yang penting, misalnya, selama aktivasi bentuk enzim yang tidak aktif, hormon protein, dan protein dari siklus mitosis. Pertumbuhan dan perkembangan sel yang normal memerlukan keseimbangan yang terkontrol dengan baik antara sintesis dan degradasi protein dan organel. Namun, dalam proses sintesis protein, karena kesalahan dalam pengoperasian peralatan sintesis, struktur molekul protein yang tidak normal, kerusakannya oleh agen kimia dan bakteri, sejumlah besar molekul yang rusak terus-menerus terbentuk. Menurut beberapa perkiraan, bagian mereka adalah sekitar sepertiga dari semua protein yang disintesis.
Sel mamalia memiliki beberapa jalur degradasi protein: melalui protease lisosom (pentida hidrolase), proteinase yang bergantung pada kalsium (endopeptidase) dan sistem proteasome. Selain itu, ada juga proteinase khusus, seperti caspases. Organel utama di mana degradasi zat dalam sel eukariotik terjadi adalah lisosom, yang mengandung banyak enzim hidrolitik. Karena proses endositosis dan berbagai jenis autophagy dalam lisosom dan fagolisosom, baik molekul protein yang rusak maupun seluruh organel dihancurkan: mitokondria yang rusak, bagian dari membran plasma, beberapa protein ekstraseluler, isi butiran sekretori.
Mekanisme penting dari degradasi protein adalah proteasome, struktur proteinase multikatalitik kompleks yang terlokalisasi di sitosol, nukleus, retikulum endoplasma, dan pada membran sel. Sistem enzim ini bertanggung jawab untuk memecah protein yang rusak serta protein sehat yang harus dikeluarkan untuk fungsi sel normal. Dalam hal ini, protein yang akan dihancurkan sebelumnya digabungkan dengan polipeptida ubiquitin spesifik. Namun, protein yang tidak ada di mana-mana juga dapat dihancurkan sebagian dalam proteasom. Pemecahan molekul protein dalam proteasom menjadi polipeptida pendek (pemrosesan) dengan presentasi berikutnya bersama-sama dengan molekul MHC tipe I merupakan tautan penting dalam penerapan kontrol imun homeostasis antigenik tubuh. Ketika fungsi proteasom melemah, akumulasi protein yang rusak dan tidak perlu terjadi, yang menyertai penuaan sel. Pelanggaran degradasi protein yang bergantung pada cyclin menyebabkan pelanggaran pembelahan sel, degradasi protein sekretori - hingga pengembangan cystofibrosis. Sebaliknya, peningkatan fungsi proteasome menyertai penipisan tubuh (AIDS, kanker).
Dengan pelanggaran degradasi protein yang ditentukan secara genetik, organisme tidak dapat hidup dan mati pada tahap awal embriogenesis. Jika pemecahan lemak atau karbohidrat terganggu, maka penyakit akumulasi (tesaurisme) terjadi. Pada saat yang sama, sejumlah besar zat atau produk tertentu dari penguraiannya yang tidak lengkap - lipid, polisakarida - terakumulasi di dalam sel, yang secara signifikan merusak fungsi sel. Paling sering diamati di epitel hati (hepatosit), neuron, fibroblas dan makrofag.
Gangguan yang didapat dalam proses disintegrasi zat dapat terjadi sebagai akibat dari proses patologis (misalnya, protein, lemak, karbohidrat dan distrofi pigmen) dan disertai dengan pembentukan zat yang tidak biasa. Pelanggaran dalam sistem proteolisis lisosom menyebabkan penurunan adaptasi selama kelaparan atau peningkatan beban, hingga terjadinya beberapa disfungsi endokrin - penurunan tingkat insulin, tiroglobulin, sitokin dan reseptornya. Pelanggaran degradasi protein memperlambat laju penyembuhan luka, menyebabkan perkembangan aterosklerosis, dan mempengaruhi respon imun. Di bawah hipoksia, perubahan pH intraseluler, cedera radiasi, ditandai dengan peningkatan peroksidasi lipid membran, serta di bawah pengaruh zat lisosomotropik - endotoksin bakteri, metabolit jamur beracun (sporofusarin), kristal silikon oksida - stabilitas membran lisosom perubahan, enzim lisosom yang diaktifkan dilepaskan ke dalam sitoplasma , yang menyebabkan penghancuran struktur sel dan kematiannya.
Bab 1
DASAR FISIOLOGI SEL
I. Dudel
membran plasma . Sel hewan dibatasi oleh membran plasma (Gbr. 1.1). Kami akan membahas strukturnya, yang sangat mirip dengan struktur banyak membran intraseluler, secara lebih rinci. Matriks utama membran terdiri dari lemak terutama fosfatidilkolin. Lipid ini terdiri dari kelompok kepala hidrofilik yang rantai hidrokarbon hidrofobik panjang melekat. Dalam air, lipid tersebut secara spontan membentuk film dua lapis setebal 4-5 nm, di mana gugus hidrofilik menghadap media berair, dan rantai hidrokarbon hidrofobik disusun dalam dua baris, membentuk fase lipid anhidrat. Membran sel adalah lipid bilayer jenis ini dan mengandung glikolipid, kolesterol, dan fosfolipid (Gbr. 1.2). Bagian hidrofilik dari glikolipid dibentuk oleh oligosakarida. Glikolipid selalu terletak di permukaan luar membran plasma, dan bagian oligosakarida dari molekul berorientasi seperti rambut terbenam di lingkungan. Tersebar di antara fosfolipid dalam jumlah molekul kolesterol yang hampir sama menstabilkan membran. Distribusi berbagai lipid di lapisan dalam dan luar membran tidak sama, bahkan di dalam lapisan yang sama terdapat daerah-daerah di mana jenis lipid tertentu terkonsentrasi. Distribusi yang tidak merata
Beras. 1.1. Gambar skema sel yang menunjukkan organel terpenting
mungkin memiliki beberapa, yang belum jelas, signifikansi fungsional.
Elemen fungsional utama yang terendam dalam matriks lipid membran yang relatif lembam adalah: tupai(Gbr. 1.2). Berat protein berkisar antara 25 hingga 75% dalam berbagai membran, tetapi karena molekul protein jauh lebih besar daripada molekul lipid, 50% beratnya setara dengan rasio 1 molekul protein dengan 50 molekul lipid. Beberapa protein menembus membran dari permukaan luar ke dalam, sementara yang lain tetap dalam satu lapisan. Molekul protein biasanya berorientasi sehingga gugus hidrofobiknya terbenam dalam membran lipid dan gugus hidrofilik polar pada permukaan membran terbenam dalam fase air. Banyak protein membran permukaan luar adalah glikoprotein; kelompok sakarida hidrofilik mereka menghadapi lingkungan ekstraseluler.
Sistem membran organel intraseluler .
Sekitar setengah dari volume sel ditempati oleh organel yang diisolasi dari sitosol oleh membran. Total permukaan membran organel intraseluler setidaknya 10 kali permukaan membran plasma. Sistem membran yang paling banyak digunakan adalah retikulum endoplasma, mewakili jaringan

Beras. 1.2.Representasi skematis dari membran plasma. Protein tertanam dalam bilayer fosfolipid, dengan beberapa merentang bilayer sementara yang lain hanya berlabuh ke lapisan luar atau dalam.
tubulus yang sangat berbelit-belit atau struktur memanjang sakular; area besar retikulum endoplasma dihiasi dengan ribosom; retikulum seperti itu disebut granular, atau kasar (Gbr. 1.1). aparatus golgi juga terdiri dari lamela terikat membran, dari mana vesikel, atau vesikel, pecah (Gbr. 1.1). Lisosom dan Peroksisom adalah vesikel khusus kecil. Dalam semua organel yang beragam ini, membran dan ruang yang dicakupnya mengandung set enzim tertentu; di dalam organel, produk metabolisme khusus menumpuk, yang digunakan untuk melakukan berbagai fungsi organel.
IntiDan mitokondria berbeda karena masing-masing organel ini dikelilingi oleh dua membran. Nukleus bertanggung jawab atas kontrol kinetik metabolisme; membran mitokondria bagian dalam yang terlipat adalah tempat metabolisme oksidatif; di sini, karena oksidasi piruvat atau asam lemak, senyawa energi tinggi adenosin trifosfat (ATP, atau ATP) disintesis.
sitoskeleton . Sitoplasma yang mengelilingi organel sama sekali tidak dapat dianggap amorf; itu diserap oleh jaringan sitoskeleton. Sitoskeleton terdiri dari mikrotubulus, filamen aktin, dan filamen menengah (Gbr. 1.1). mikrotubulus memiliki diameter luar sekitar 25 nm; mereka terbentuk, seperti polimer normal, sebagai hasil dari perakitan molekul protein tubulin. filamen aktin- serat kontraktil yang terletak di lapisan dekat membran dan di seluruh sel - terutama mengambil bagian dalam proses yang terkait dengan gerakan. Filamen perantara terdiri dari blok komposisi kimia yang berbeda dalam berbagai jenis sel; mereka membentuk berbagai hubungan antara dua elemen lain dari sitoskeleton yang disebutkan di atas. Organel dan membran plasma juga berhubungan dengan sitoskeleton, yang tidak hanya mempertahankan bentuk sel dan posisi organel di dalamnya, tetapi juga menentukan perubahan bentuk sel dan mobilitasnya.
sitosol . Sekitar setengah dari volume sel ditempati oleh sitosol. Karena proteinnya kira-kira 20% (berdasarkan berat), ini lebih berupa gel daripada larutan berair. Molekul kecil, termasuk organik dan anorganik ion, terlarut dalam fase air. Antara sel dan lingkungan (ruang ekstraseluler) terjadi pertukaran ion; proses pertukaran ini akan dibahas di bagian selanjutnya. Konsentrasi ion di ruang ekstraseluler dipertahankan dengan akurasi yang cukup besar pada tingkat yang konstan; konsentrasi intraseluler masing-masing ion juga memiliki tingkat tertentu yang berbeda dari yang di luar sel (Tabel 1.1). Kation yang paling umum di lingkungan ekstraseluler adalah Na+ dalam sel, konsentrasinya lebih dari 10 kali lebih rendah. Sebaliknya, di dalam sel konsentrasi K + adalah yang tertinggi, di luar sel lebih rendah lebih dari satu urutan besarnya. Gradien terbesar antara konsentrasi ekstraseluler dan intraseluler ada untuk Ca 2+, konsentrasi ion bebas yang di dalam sel setidaknya 10.000 kali lebih rendah daripada di luarnya. Tidak semua ion terlarut dalam sitosol; beberapa di antaranya diadsorpsi pada protein atau disimpan dalam organel. Misalnya, dalam kasus Ca2+ ion terikat jauh lebih banyak daripada yang bebas. Sebagian besar protein sitosol adalah enzim, dengan partisipasi yang melakukan banyak proses metabolisme antara: glikolisis dan glukoneogenesis, sintesis atau penghancuran asam amino, sintesis protein pada ribosom (Gbr. 1.1). Sitosol juga mengandung tetesan lipid dan butiran glikogen, yang berfungsi sebagai cadangan molekul penting.
Tabel 1.1.Konsentrasi intra dan ekstraseluler ion dalam sel otot hewan homoiothermic. TETAPI – – “anion seluler dengan berat molekul tinggi”
|
Konsentrasi intraseluler |
Konsentrasi ekstraseluler |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca2+ |
10 –7 10 –8 mmol/l |
Ca2+ |
2 mmol/l |
|
Dengan aku - |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO3 - |
8 mmol/l |
HCO3 - |
27 mmol/l |
|
SEBUAH- |
155 mmol/l |
Lainnya kation |
5 mmol/l |
|
Potensi istirahat -90 mV |
|||
1.2. Pertukaran zat antara sel dan lingkungan
Kami telah menjelaskan secara singkat struktur sel untuk menggunakan deskripsi ini untuk meninjau dasar-dasar fisiologi sel. Dalam kasus apa pun sel tidak dapat dianggap sebagai formasi statis, karena ada pertukaran zat yang konstan antara berbagai kompartemen intraseluler, serta antara kompartemen dan lingkungan. Struktur sel berada dalam keseimbangan dinamis, dan interaksi sel satu sama lain dan dengan lingkungan eksternal merupakan kondisi yang diperlukan untuk mempertahankan kehidupan organisme yang berfungsi. Dalam bab ini, kita akan mempertimbangkan mekanisme fundamental dari pertukaran semacam itu. Dalam bab-bab selanjutnya, mekanisme-mekanisme ini akan dibahas dalam kaitannya dengan sel saraf dan fungsinya;
namun, mekanisme yang sama mendasari fungsi semua organ lainnya.
Difusi.Proses paling sederhana untuk memindahkan suatu zat adalah difusi. Dalam larutan (atau gas), atom dan molekul bergerak bebas, dan perbedaan konsentrasi diseimbangkan dengan difusi. Pertimbangkan dua volume yang diisi dengan cairan atau gas (Gbr. 1.3), di mana zat memiliki konsentrasi c1 dan c2 dan dipisahkan oleh lapisan dengan luas permukaan A dan tebal D. Aliran materi m dalam waktu t dijelaskan Hukum difusi pertama Fick:
dm/ dt= DSEBUAH/ D ( C 1 –С 2)=DSEBUAH/ DD C(1)
di mana D adalah koefisien difusi, yang konstan untuk zat, pelarut, dan suhu tertentu. Dalam bentuk yang lebih umum, untuk perbedaan konsentrasi dc pada jarak dx
dm/dt= -D A dc/dx,(2)
aliran melalui bagian A sebanding dengan gradien konsentrasi dc/dx . Tanda minus muncul dalam persamaan karena perubahan konsentrasi dalam arah x adalah negatif.
Difusi adalah proses yang paling penting dimana sebagian besar molekul dalam larutan air bergerak dalam jarak pendek. Ini juga berlaku untuk pergerakan mereka di dalam sel sejauh difusi tidak dihalangi oleh membran. Banyak zat dapat dengan bebas berdifusi melalui membran lipid, terutama air dan gas terlarut seperti O 2 dan CO 2 . larut dalam lemak

Beras. 1.3.Skema kuantitatif difusi. Kedua ruang dipisahkan oleh lapisan ketebalanDdan daerah TETAPI. C; - konsentrasi partikel tinggi di bagian kiri volume, C :, - konsentrasi partikel rendah di bagian kanan bagian, permukaan merah muda adalah gradien konsentrasi pada lapisan difusi. Fluks difusi dm/dt – lihat persamaan (1)
zat juga berdifusi dengan baik melalui membran; ini juga berlaku untuk molekul polar yang relatif kecil, seperti etanol dan urea, sementara gula melewati lapisan lipid dengan susah payah. Pada saat yang sama, lapisan lipid praktis tidak dapat ditembus oleh molekul bermuatan, termasuk bahkan ion anorganik. Untuk non-elektrolit, persamaan difusi (1) biasanya diubah dengan menggabungkan karakteristik membran dan zat yang berdifusi menjadi satu. parameter-permeabilitas (P):
dm/dt=P AD C.(3)
pada gambar. 1,4 dibandingkan permeabilitas (P) membran lipid untuk berbagai molekul.
Difusi melalui pori-pori membran . Membran plasma (dan membran sel lainnya) tidak hanya permeabel terhadap zat yang berdifusi melalui lapisan lipid, tetapi juga terhadap banyak ion, gula, asam amino, dan nukleotida. Zat-zat ini melintasi membran melalui pori-pori yang dibentuk oleh protein transportasi, tertanam dalam membran. Di dalam protein seperti itu, ada saluran berisi air dengan diameter kurang dari 1 nm di mana molekul kecil dapat berdifusi. Mereka bergerak sepanjang gradien konsentrasi, dan jika mereka membawa muatan, maka pergerakan mereka melalui saluran juga diatur oleh potensial membran. Saluran membran relatif selektif

Beras. 1.4.Permeabilitas lapisan ganda lipid buatan untuk berbagai zat
sehubungan dengan jenis molekul yang dapat melewatinya, misalnya, ada saluran kalium, natrium, dan kalsium, yang masing-masing tidak dapat ditembus oleh hampir semua ion kecuali satu ion tertentu. Seperti selektivitas karena muatan atau struktur situs pengikatan di dinding saluran, yang memfasilitasi pengangkutan molekul tertentu dan mencegah penetrasi zat lain melalui saluran (Gbr. 1,5, A) .
Dibalik perilaku saluran ion membran Sangat mudah untuk mengamati, karena arus yang timbul selama pergerakan ion dapat diukur, dan bahkan untuk satu saluran. Ditunjukkan bahwa saluran secara spontan dan dengan frekuensi tinggi mengubah keadaannya dari terbuka menjadi tertutup. Saluran kalium dicirikan oleh pulsa arus dengan amplitudo sekitar 2 pA (2 10 -12 A) dan durasi beberapa milidetik (lihat Gambar 2.12, hal. 37) [3]. Selama periode ini, puluhan ribu ion melewatinya. Transisi protein dari satu konformasi ke yang lain dipelajari dengan difraksi sinar-X, spektroskopi Mössbauer dan resonansi magnetik nuklir (NMR). Protein dengan demikian sangat dinamis, struktur bergerak, dan saluran melalui protein bukan hanya tabung kaku berisi air (Gambar 1.5A) tetapi labirin kelompok molekul dan muatan yang bergerak cepat. Tanggapan dinamis saluran ini tercermin dalam profil energi saluran, ditunjukkan pada gambar. 1.5, B. Di sini, absis menunjukkan panjang saluran dari larutan eksternal dengan konsentrasi ion C 0 dan potensial 0 ke larutan internal dengan konsentrasi C 1 dan potensial E. Sumbu y

Beras. 1.5.A. Skema protein yang membentuk saluran kalium yang tertanam dalam lapisan ganda lipid membran plasma. Empat muatan negatif dipasang di "dinding" saluran. B. Profil energi skema saluran yang ditunjukkan pada gambar. A. Pada sumbu y, nilai-nilai energi kinetik yang diperlukan untuk melewati saluran diplot; sepanjang sumbu absis, jarak antara permukaan dalam dan luar membran. Energi minimum sesuai dengan situs pengikatan ion bermuatan positif dengan muatan negatif tetap di dinding saluran. Energi maxima sesuai dengan hambatan difusi di saluran. Diasumsikan bahwa konformasi protein saluran berosilasi secara spontan; opsi untuk profil energi ditunjukkan oleh garis padat dan putus-putus; osilasi ini sangat memudahkan pengikatan ion saat mengatasi penghalang energi (tetapi dengan perubahan)
tingkat energi ion di situs pengikatan saluran ditunjukkan; puncak dalam grafik mewakili penghalang permeabilitas yang harus diatasi oleh energi ion untuk menembus saluran, dan "penurunan" grafik mewakili keadaan yang relatif stabil (mengikat). Terlepas dari hambatan puncak energi, ion dapat menembus saluran jika profil energi berputar secara spontan; ion, dengan demikian, dapat tiba-tiba muncul "di sisi lain" dari puncak energi dan dapat terus bergerak ke dalam sel. Bergantung pada muatan, ukuran, dan derajat hidrasi ion dan kemampuannya untuk berikatan dengan struktur dinding saluran, profil energi saluran bervariasi untuk ion yang berbeda, yang dapat menjelaskan selektivitas masing-masing jenis saluran.
Kesetimbangan difusi ion . Difusi berbagai ion melalui saluran membran harus mengarah pada penghapusan perbedaan konsentrasi antara lingkungan ekstra dan intraseluler. Namun, seperti yang dapat dilihat dari Tabel. 1.1, perbedaan seperti itu tetap ada, jadi pasti ada keseimbangan antara difusi dan proses transpor lainnya melintasi membran. Dua bagian berikutnya membahas cara-cara di mana keseimbangan semacam itu terbentuk. Dalam kasus ion, kesetimbangan difusi dipengaruhi oleh muatannya. Difusi molekul tidak bermuatan disediakan oleh perbedaan konsentrasi dc , dan ketika konsentrasinya sama, transportasi yang sebenarnya berhenti. Partikel bermuatan juga dipengaruhi oleh medan listrik. Misalnya, ketika ion kalium keluar dari sel sepanjang gradien konsentrasinya, ia membawa satu muatan positif. Dengan demikian, lingkungan intraseluler menjadi lebih bermuatan negatif, menghasilkan perbedaan potensial melintasi membran. Muatan negatif intraselular mencegah ion kalium baru meninggalkan sel, dan ion-ion yang meninggalkan sel lebih lanjut akan meningkatkan muatan pada membran. Aliran ion kalium berhenti ketika aksi medan listrik mengkompensasi tekanan difusi karena perbedaan konsentrasi. Ion terus melewati membran, tetapi dalam jumlah yang sama di kedua arah. Oleh karena itu, untuk perbedaan konsentrasi ion tertentu pada membran, terdapat potensial keseimbangan E ion di mana aliran ion melalui membran berhenti. Potensial kesetimbangan dapat dengan mudah ditentukan dengan menggunakan Persamaan Nernst:
Eion= RT/ zF* lnKeluar/ C dalam(4)
dimana R adalah konstanta gas, T adalah suhu absolut, z adalah valensi ion (negatif untuk anion) Keluar adalah konsentrasi ion ekstraseluler, C dalam adalah konsentrasi ion intraseluler, F nomor Faraday. Jika kita memasukkan konstanta ke dalam persamaan, maka pada suhu tubuh (T = 310 K) potensial kesetimbangan untuk ion kalium E K sama dengan:
ek= –61 MB log /(5)
Jika [ K + keluar ]/[ K + masuk ] = 39, sebagai berikut dari Tabel. 1.1, maka
Ek= -61 m B log 39= -97 mV.
Memang, ditemukan bahwa semua sel memiliki potensial membran; dalam sel otot mamalia, levelnya sekitar -90 mV. Tergantung pada kondisi dan konsentrasi relatif ion, sel dapat memiliki potensial membran berkisar antara -40 hingga -120mV. Untuk sel pada contoh di atas (Tabel 1.1) potensi istirahat, sama dengan sekitar -90 mV, menunjukkan bahwa fluks ion kalium melalui saluran membran kira-kira dalam kesetimbangan. Ini tidak mengherankan, karena keadaan terbuka saluran kalium di membran istirahat adalah yang paling mungkin; membran paling permeabel terhadap ion kalium. Potensi membran, bagaimanapun, ditentukan oleh fluks ion lain juga.
Kemudahan partikel tak bermuatan dapat berdifusi melalui membran dikuantifikasi dalam persamaan (3). Permeabilitas terhadap partikel bermuatan dijelaskan oleh persamaan yang sedikit lebih kompleks:
P= M RT/ dF(6)
di mana Madalah mobilitas ion dalam membran, D - Ketebalan membran, a R , T dan F konstanta termodinamika yang diketahui. Nilai permeabilitas untuk berbagai ion yang ditentukan dengan cara ini dapat digunakan untuk menghitung potensial membran em ketika ion kalium, natrium dan klorida melewati membran secara bersamaan (dengan permeabilitas P K, P Na dan P Cl masing-masing). Diasumsikan bahwa potensial turun secara merata di dalam membran, sehingga kuat medannya konstan. Dalam hal ini, itu berlaku Persamaan Goldman, atau persamaan medan konstan :
Em= R T/ F * ln(P K + P Na + P Cl )/ (PK + P Na + P Cl )(7)
Untuk sebagian besar membran sel P K sekitar 30 kali lebih tinggi dari R tidak (lihat juga bagian 1.3). Nilai relatif PCl sangat bervariasi; untuk banyak membran PCl kecil dibandingkan dengan R K , namun untuk orang lain (misalnya di otot rangka) PCl , jauh lebih tinggi dari R K
Transpor aktif, pompa natrium . Bagian sebelumnya menjelaskan difusi pasif ion dan potensial membran yang dihasilkan pada konsentrasi ion intra dan ekstraseluler tertentu. Namun, sebagai hasil dari proses ini, konsentrasi ion di dalam sel tidak secara otomatis distabilkan, karena membran
potensial sedikit lebih elektronegatif daripada E K , dan masih banyak lagi dibandingkan dengan E tidak (sekitar +60 mV). Karena difusi, konsentrasi ion intraseluler, setidaknya kalium dan natrium, harus sama dengan konsentrasi ekstraseluler. Stabilitas gradien ion dicapai melalui transpor aktif: protein membran mengangkut ion melintasi membran melawan gradien konsentrasi listrik dan (atau), mengkonsumsi energi metabolik untuk ini. Proses transpor aktif yang paling penting adalah kerja Na/K - pompa yang ada di hampir semua sel;
pompa memompa ion natrium keluar sel sekaligus memompa ion kalium ke dalam sel. Hal ini memastikan konsentrasi ion natrium intraseluler yang rendah dan kalium yang tinggi (Tabel 1.1). Gradien konsentrasi ion natrium pada membran memiliki fungsi spesifik yang terkait dengan transmisi informasi dalam bentuk impuls listrik (lihat Bagian 2.2), serta dengan pemeliharaan mekanisme transpor aktif lainnya dan regulasi volume sel (lihat di bawah). Oleh karena itu, tidak mengherankan bahwa lebih dari 1/3 energi yang dikonsumsi oleh sel dihabiskan untuk pompa Na / K, dan di beberapa sel yang paling aktif hingga 70% energi dihabiskan untuk operasinya.
Protein transpor Na/K adalah ATPase. Pada permukaan bagian dalam membran, ia memecah ATP menjadi ADP dan fosfat (Gbr. 1.6). Energi dari satu molekul ATP digunakan untuk mengangkut tiga ion natrium dari sel dan secara bersamaan dua ion kalium ke dalam sel, yaitu, secara total, satu muatan positif dikeluarkan dari sel dalam satu siklus. Jadi pompa Na/K adalah elektrogenik(menciptakan arus listrik melalui membran), yang menyebabkan peningkatan elektronegativitas potensial membran sekitar 10 mV. Protein transpor melakukan operasi ini dengan kecepatan tinggi: dari 150 hingga 600 ion natrium per detik. Urutan asam amino dari protein transpor diketahui, tetapi mekanisme transpor pertukaran kompleks ini belum jelas. Proses ini dijelaskan dengan menggunakan profil energi dari transfer ion natrium atau kalium oleh protein (Gbr. 1.5.5). Berdasarkan sifat perubahan profil ini, terkait dengan perubahan konstan dalam konformasi protein transpor (proses yang membutuhkan energi), seseorang dapat menilai stoikiometri pertukaran: dua ion kalium ditukar dengan tiga ion natrium.
Pompa Na/K, seperti terisolasi Na+ /K + - ATPase membran yang bergantung, secara khusus dihambat oleh ouabain glikosida jantung (strophanthin). Karena pengoperasian pompa Na/K merupakan reaksi kimia multi-tahap, maka, seperti semua reaksi kimia, sebagian besar bergantung pada suhu, yang

Beras. 1.6.Skema Na/K-pompa-ATPase (terbenam dalam lipid bilayer membran plasma), yang dalam satu siklus mengeluarkan tiga ion Na + dari sel melawan gradien potensial dan konsentrasi dan membawa dua ion K ke dalam sel + . Selama proses ini, satu molekul ATP dipecah menjadi ADP dan fosfat. Dalam diagram, ATPase ditampilkan sebagai dimer yang terdiri dari subunit besar (fungsional) dan kecil; dalam membran itu ada sebagai tetramer yang dibentuk oleh dua subunit besar dan dua kecil
ditunjukkan pada gambar. 1.7. Di sini aliran ion natrium dari sel otot ditunjukkan dalam kaitannya dengan waktu; ini secara praktis setara dengan aliran ion natrium yang diperantarai oleh operasi pompa Na/K, karena aliran pasif ion natrium melawan gradien konsentrasi dan potensial sangat kecil. Jika sediaan didinginkan sekitar 18°C, maka aliran ion natrium dari sel akan berkurang dengan cepat sebanyak 15 kali, dan segera setelah dipanaskan akan dikembalikan ke tingkat semula. Penurunan aliran ion natrium dari sel seperti itu beberapa kali lebih besar daripada yang sesuai dengan ketergantungan suhu dari proses difusi atau reaksi kimia sederhana. Efek serupa diamati ketika energi metabolik terkuras akibat keracunan dinitrofenol (DNP) (Gbr. 1.7.5). Oleh karena itu, aliran ion natrium dari sel disediakan oleh reaksi yang bergantung pada energi - pompa aktif. Karakteristik lain dari pompa, bersama dengan ketergantungan suhu dan energi yang signifikan, adalah adanya tingkat kejenuhan (seperti semua reaksi kimia lainnya); ini berarti bahwa kecepatan pompa tidak dapat meningkat tanpa batas karena konsentrasi ion yang diangkut meningkat (Gbr. 1.8). Sebaliknya, aliran zat yang berdifusi secara pasif tumbuh sebanding dengan perbedaan konsentrasi sesuai dengan hukum difusi (persamaan 1 dan 2).

Beras. 1.7. A, B Transpor aktif + . sumbu Y: aliran radioaktif 24 Na + dari sel (imp./min). sumbu absis: waktu sejak awal percobaan. TETAPI. Sel didinginkan dari 18,3°C menjadi 0,5°C; mengalir Na+ keluar dari sel selama periode ini terhambat. B. Penekanan aliran Na + dari sel dengan dinitrofenol (DNF) pada konsentrasi 0,2 mmol/l (sebagaimana telah diubah)
Selain pompa Na / K, membran plasma mengandung setidaknya satu pompa lagi - kalsium; pompa ini memompa keluar ion kalsium (Ca 2+) dari sel dan terlibat dalam mempertahankan konsentrasi intraseluler mereka pada tingkat yang sangat rendah (Tabel 1.1). Pompa kalsium hadir pada kepadatan yang sangat tinggi di retikulum sarkoplasma sel otot, yang mengakumulasi ion kalsium sebagai akibat pemecahan molekul ATP (lihat Bab 4).
Efek pompa Na/K pada potensial membran dan volume sel . pada gambar. 1.9 menunjukkan berbagai komponen arus membran dan menunjukkan konsentrasi ion intraseluler yang

Beras. 1.8.Rasio antara laju pengangkutan molekul dan konsentrasinya (pada titik masuk ke saluran atau pada titik pengikatan pompa) selama difusi melalui saluran atau selama pengangkutan pemompaan. Yang terakhir jenuh pada konsentrasi tinggi (laju maksimum, Vmax ) nilai pada absis yang sesuai dengan setengah dari kecepatan pompa maksimum ( Vmax /2), adalah konsentrasi kesetimbangan KE M

Beras. 1.9.Diagram yang menunjukkan konsentrasi Na+ , K+ dan Cl- di dalam dan di luar sel dan cara ion-ion ini menembus membran sel (melalui saluran ion tertentu atau dengan bantuan pompa Na/K. Dengan gradien konsentrasi yang diberikan, potensial kesetimbangan E Na, E K dan E C l - sama dengan yang ditunjukkan, potensial membran em = – 90 mV
memastikan keberadaan mereka. Arus keluar ion kalium diamati melalui saluran kalium, karena potensial membran agak lebih elektropositif daripada potensial kesetimbangan untuk ion kalium. Konduktivitas total saluran natrium jauh lebih rendah daripada saluran kalium; saluran natrium terbuka lebih jarang daripada saluran kalium pada potensial istirahat; namun, kira-kira jumlah ion natrium yang masuk ke dalam sel sama dengan jumlah ion kalium yang keluar, karena konsentrasi yang besar dan gradien potensial diperlukan untuk difusi ion natrium ke dalam sel. Pompa Na/K memberikan kompensasi yang ideal untuk arus difusi pasif, karena ia mengangkut ion natrium keluar dari sel dan ion kalium ke dalamnya. Dengan demikian, pompa bersifat elektrogenik karena perbedaan jumlah muatan yang ditransfer ke dalam dan ke luar sel, yang pada kecepatan normal operasinya, menciptakan potensial membran sekitar 10 mV lebih elektronegatif daripada jika dihasilkan oleh arus ion pasif saja (lihat Persamaan 7). Akibatnya, potensial membran mendekati potensial kesetimbangan kalium, yang mengurangi kebocoran ion kalium. Tidak ada aktivitas/K-pompa diatur oleh konsentrasi ion natrium intraseluler. Kecepatan pompa melambat karena konsentrasi ion natrium yang akan dikeluarkan dari sel menurun (Gbr. 1.8), sehingga operasi pompa dan aliran ion natrium ke dalam sel saling menyeimbangkan, mempertahankan konsentrasi intraseluler ion natrium pada tingkat sekitar 10 mmol / l.
Untuk menjaga keseimbangan antara pemompaan dan arus membran pasif, dibutuhkan lebih banyak molekul pompa Na/K daripada protein saluran untuk ion kalium dan natrium. Ketika saluran terbuka, puluhan ribu ion melewatinya dalam beberapa milidetik (lihat di atas), dan karena saluran biasanya dibuka beberapa kali per detik, lebih dari 105 ion melewatinya secara total selama waktu ini. Sebuah protein pompa tunggal menggerakkan beberapa ratus ion natrium per detik, sehingga membran plasma harus mengandung sekitar 1000 kali lebih banyak molekul pompa daripada molekul saluran. Pengukuran arus saluran saat istirahat menunjukkan rata-rata satu kalium dan satu saluran terbuka natrium per 1 m 2 membran; maka dari sini sekitar 1000 molekul pompa Na/K harus ada di ruang yang sama, yaitu. jarak antara mereka rata-rata 34 nm; diameter protein pemompaan sebagai protein saluran adalah 8-10 nm. Dengan demikian, membran cukup jenuh dengan molekul pemompa
Fakta bahwa aliran ion natrium ke dalam sel, dan ion kalium keluar sel dikompensasi oleh pengoperasian pompa, memiliki konsekuensi lain, yang terdiri dari mempertahankan tekanan osmotik yang stabil dan volume konstan. Di dalam sel terdapat konsentrasi tinggi anion besar, terutama protein (A - pada Tabel 1.1), yang tidak mampu menembus membran (atau menembusnya dengan sangat lambat) dan oleh karena itu merupakan komponen tetap di dalam sel. Untuk menyeimbangkan muatan anion ini, diperlukan jumlah kation yang sama. Karena aksi pompa Na/K, kation ini terutama ion kalium. Peningkatan yang signifikan dalam konsentrasi ion intraseluler dapat terjadi hanya dengan peningkatan konsentrasi anion karena aliran C1 - sepanjang gradien konsentrasi ke dalam sel (Tabel 1.1), tetapi potensial membran melawan hal ini. Arus masuk Cl- diamati hanya sampai potensial kesetimbangan untuk ion klorida tercapai; ini diamati ketika gradien ion klorida hampir berlawanan dengan gradien ion kalium, karena ion klorida bermuatan negatif (persamaan 4). Dengan demikian, konsentrasi ion klorida intraseluler yang rendah terbentuk, sesuai dengan konsentrasi ion kalium ekstraseluler yang rendah. Hasilnya adalah pembatasan jumlah total ion dalam sel. Jika potensial membran turun ketika pompa Na/K diblokir, misalnya selama anoksia, maka potensial kesetimbangan untuk ion klorida menurun, dan konsentrasi ion klorida intraseluler meningkat. Mengembalikan keseimbangan muatan, ion kalium juga masuk ke dalam sel; konsentrasi total ion dalam sel meningkat, yang meningkatkan tekanan osmotik; ini memaksa air untuk masuk ke dalam sel. Sel membengkak. Pembengkakan ini terlihat in vivo dalam kondisi kekurangan energi.
gradien konsentrasi tidak + sebagai kekuatan pendorong untuk transportasi membran . Pentingnya pompa Na/K-untuk sel tidak terbatas pada stabilisasi gradien K+ dan Na+ normal melintasi membran. Energi yang tersimpan dalam gradien membran Na+ , sering digunakan untuk menyediakan transportasi membran untuk zat lain. Misalnya, pada gambar. 1.10 menunjukkan "simport" Na+ dan molekul gula ke dalam sel. Protein transpor membran membawa molekul gula ke dalam sel bahkan melawan gradien konsentrasi, sementara Na + bergerak sepanjang gradien konsentrasi dan potensial, menyediakan energi untuk transportasi gula. Pengangkutan gula semacam itu sepenuhnya bergantung pada keberadaan gradien yang tinggi Na+ ; jika konsentrasi intraseluler Na+ meningkat secara signifikan, maka pengangkutan gula berhenti. Untuk berbagai C Akharov, ada sistem symport yang berbeda. Transportasi asam amino dalam sangkar yang mirip dengan transportasi C akharov ditunjukkan pada Gambar. 1.10; itu juga disediakan oleh gradien Na+ , setidaknya ada lima sistem symport yang berbeda, masing-masing khusus untuk satu kelompok asam amino terkait.
Selain sistem symport, ada juga "antiport". Salah satunya, misalnya, mentransfer satu ion kalsium keluar sel dalam satu siklus sebagai ganti tiga ion natrium yang masuk (Gbr. 1.10). Energi untuk transportasi Ca2+ terbentuk karena masuknya tiga ion natrium sepanjang gradien konsentrasi dan potensial. Energi ini cukup (pada potensial istirahat) untuk mempertahankan gradien ion kalsium yang tinggi (dari kurang dari 10-7 mol/l di dalam sel hingga sekitar 2 mmol/l di luar sel).
Endo- dan eksositosis . Untuk zat tertentu yang masuk ke dalam sel atau harus dikeluarkan

Beras. 1.10.Protein tertanam dalam lipid bilayer membran memediasi impor glukosa dan Na + ke dalam sel, serta Ca2+/Na+ -antiport, di mana kekuatan pendorongnya adalah gradien Na + pada membran sel
dari sana, tidak ada saluran transportasi; zat tersebut termasuk, misalnya, protein dan kolesterol. Mereka dapat melewati membran plasma ke dalam vesikel atau gelembung, melalui endositosis dan eksositosis. pada gambar. 1.11 menunjukkan mekanisme utama dari proses ini. Selama eksositosis, organel tertentu (lihat di bawah) membentuk vesikel berisi zat yang perlu dikeluarkan dari sel, seperti hormon atau enzim ekstraseluler. Ketika vesikel tersebut mencapai membran plasma, membran lipidnya menyatu dengannya, sehingga memungkinkan isinya keluar ke lingkungan eksternal. Dalam proses yang berlawanan, endositosis, membran plasma berinvaginasi, membentuk lubang, yang kemudian memperdalam dan menutup, membentuk vesikel intraseluler yang diisi dengan cairan ekstraseluler dan beberapa makromolekul. Untuk memastikan fusi membran dan penutupan vesikel ini, elemen kontraktil sitoskeleton bekerja bersama dengan membran itu sendiri (lihat di bawah). Endositosis tidak selalu melibatkan hanya menangkap media ekstraseluler ke dalam sel. Membran sel mengandung, sering diatur ke dalam kelompok khusus, reseptor spesifik untuk makromolekul seperti insulin atau antigen. Setelah makromolekul ini berikatan dengan reseptornya, endositosis terjadi di daerah membran yang mengelilingi reseptor, dan makromolekul tersebut secara selektif diangkut ke dalam sel (Gbr. 1.12, B).
Endo- dan eksositosis terjadi terus menerus dalam sel. Jumlah bahan membran yang bersirkulasi signifikan; dalam waktu 1 jam, makrofag menyerap dalam bentuk vesikel dua kali luas permukaan membran sitoplasmanya. Pada kebanyakan sel, pergantian bahan membran tidak begitu intensif, tetapi tetap harus signifikan.

Beras. 1.11.eksositosis dan endositosis. Ke atas: vesikel intraseluler menyatu dengan lapisan ganda lipid dari membran plasma dan membuka ke dalam ruang ekstraseluler. Proses ini disebut eksositosis. Di dasar: membran plasma berinvaginasi di area kecil dan mengikat vesikel yang berisi bahan ekstraseluler. Proses ini disebut endositosis.
1.3. Transportasi zat di dalam sel
Endo- dan eksositosis tidak hanya proses pengangkutan zat melalui membran sel, tetapi juga proses pertukaran membran - komponen struktural sel itu sendiri. Subjek pertimbangan dalam bagian ini adalah proses transpor serupa lainnya di dalam sel dan organelnya.

Beras. 1.12. A-B. Skema proses, termasuk ekso-dan endositosis. TETAPI. Protein yang disintesis dalam retikulum endoplasma granular diangkut melalui aparatus Golgi ke membran plasma, di mana ia disekresikan oleh eksositosis. B. Kolesterol terikat pada LDL (low density lipoprotein) partikel menempel pada membran plasma, menginduksi pembentukan vesikel endositik di wilayah membran, dan diangkut ke lisosom, di mana ia dilepaskan. DI DALAM. Materi ekstraseluler ditangkap selama endositosis (pada gambar di kanan), diangkut melalui sel dalam vesikel, atau vesikel, dan dilepaskan melalui eksositosis (pada gambar kiri)
Difusi . Secara alami, dalam sitosol, perbedaan konsentrasi dihilangkan dengan difusi; hal yang sama berlaku untuk cairan yang terkandung dalam organel. Karena konsentrasi protein terlarut yang tinggi, difusi di sini jauh lebih lambat daripada di air. Membran lipid—di sekitar sel dan di dalam organel—adalah cairan dua dimensi tempat terjadinya difusi. Lipid pada membran bilayer berdifusi di dalam lapisannya sendiri, jarang berpindah dari satu ke yang lain. Protein yang terbenam di dalamnya juga cukup mobile; mereka berputar pada sumbu yang tegak lurus terhadap membran atau berdifusi secara lateral dengan konstanta difusi yang sangat berbeda, 2–10.000 kali lebih lambat dari fosfolipid. Jadi, jika beberapa protein bergerak bebas di lapisan lipid dan pada kecepatan yang sama dengan molekul lipid itu sendiri, maka yang lain akan ditambatkan, mis. cukup kuat terkait dengan sitoskeleton. Ada agregat "permanen" dari protein spesifik dalam membran, seperti struktur sel saraf pra dan pascasinaps. Protein yang bergerak bebas dapat ditunjukkan dengan mengikatnya ke pewarna fluoresen, yang diinduksi untuk bersinar dengan menerangi area kecil membran dengan kilatan singkat. Eksperimen semacam itu menunjukkan bahwa dalam waktu kurang dari 1 menit, protein yang terikat pada pewarna didistribusikan secara merata di atas membran pada jarak hingga 10 m.
Transpor aktif dalam membran organel .
Proses transpor aktif, yang memainkan peran penting dalam fungsi membran plasma, juga terjadi di dalam sel, di membran organel. Isi spesifik dari berbagai organel dibuat sebagian oleh sintesis internal dan sebagian oleh transpor aktif dari sitosol. Salah satu contoh yang terakhir adalah pompa Ca2+ yang disebutkan di atas dalam retikulum sarkoplasma sel otot. Sangat menarik bahwa dalam kasus sintesis ATP di mitokondria, prinsip yang berlawanan berlaku untuk apa yang terjadi di pompa ATPase dari membran plasma (Gbr. 1.6). Selama sintesis ATP, metabolisme oksidatif mengarah pada pembentukan gradien yang curam H+ pada membran dalam. Gradien ini adalah kekuatan pendorong untuk proses yang berlawanan dengan siklus pemompaan transpor molekul aktif: ion H + bergerak melintasi membran sepanjang gradien, dan energi yang dilepaskan sebagai hasilnya menyediakan sintesis ATP dari ADP dan fosfat . ATP yang dihasilkan, pada gilirannya, menyediakan energi ke sel, termasuk untuk transpor aktif.
Transportasi dalam vesikel . Sel memiliki sejumlah besar organel dan vesikel terkait (Gbr. 1.1). Organel-organel ini, dan terutama vesikel, bergerak konstan, mengangkut isinya ke organel lain atau ke membran plasma. Vesikel juga dapat bermigrasi dari membran sel ke organel, seperti pada endositosis.
Proses sekresi protein ditunjukkan pada gambar. 1.12 TETAPI. Protein disintesis di dekat inti sel pada ribosom yang terkait dengan retikulum endoplasma (yang disebut retikulum endoplasma granular, atau kasar); Begitu berada di retikulum endoplasma, protein dikemas ke dalam vesikel transpor, yang dipisahkan dari organel dan bermigrasi ke aparatus Golgi. Di sini mereka bergabung dengan tangki aparatus Golgi, di mana protein dimodifikasi (yaitu, diubah menjadi glikoprotein). Di ujung tangki, vesikel terpisah lagi. Vesikel sekretori yang membawa protein yang dimodifikasi bergerak menuju membran plasma dan melepaskan isinya melalui eksositosis.
Contoh lain dari jalur transportasi dalam sel ditunjukkan pada Gambar. 1.12, B; adalah pengambilan kolesterol oleh sel. Kolesterol yang diangkut dalam darah terutama terkait dengan protein, seperti partikel "lipoprotein densitas rendah"(LNP). Partikel-partikel ini menempel pada situs reseptor LDL spesifik pada membran di mana endositosis terjadi dan LDL diangkut ke dalam sel dalam vesikel "dilapisi". Vesikel ini menyatu untuk membentuk endosom dan kehilangan 'dering' selama proses ini. Endosom pada gilirannya menyatu dengan lisosom primer yang mengandung sebagian besar enzim hidrolitik untuk membentuk lisosom sekunder yang lebih besar. Di dalamnya, kolesterol dilepaskan dari partikel LDL dan berdifusi ke dalam sitosol, di mana ia tersedia, misalnya, untuk sintesis membran lipid. Vesikel yang tidak mengandung LDL juga dipisahkan dari endosom, yang bergerak secara khusus ke membran plasma dan bergabung dengannya, mengembalikan bahan membran dan, mungkin, reseptor LDL. Dari saat partikel LDL mengikat membran, 10-15 menit berlalu sampai kolesterol dilepaskan dari lisosom sekunder. Gangguan dalam pengikatan dan penyerapan LDL, yaitu pasokan kolesterol ke sel, memainkan peran yang menentukan dalam perkembangan penyakit yang serius dan meluas, aterosklerosis ("pengerasan" arteri).
Ada banyak rute transportasi lain yang serupa dengan yang ditunjukkan pada Gambar. 1.11 dan 1.12,A, dengan bantuan vesikel spesifik yang bergerak di dalam sel. Tidak diketahui secara pasti bagaimana mereka bergerak, tetapi elemen sitoskeleton mungkin terlibat dalam proses ini. Vesikel dapat meluncur di sepanjang mikrotubulus, dalam hal ini energi untuk gerakan tampaknya disediakan oleh protein terkait vesikel, ATPase (lihat di bawah). Tetap tidak dapat dipahami berapa banyak vesikel yang berbeda, bergerak satu demi satu ke segala arah, sampai ke tujuan mereka. Mereka jelas perlu "ditandai" sedemikian rupa sehingga dikenali oleh sistem transportasi dan diubah menjadi gerakan yang bertujuan.
Transportasi dengan pembentukan dan penghancuran organel . Sampai saat ini, kami telah menganggap endo- dan eksositosis sebagai proses pengangkutan isi vesikel. Ada aspek lain dari proses ini, yang terdiri dari fakta bahwa penghapusan langsung membran plasma di satu area permukaan sel oleh endositosis dan, sebaliknya, penambahannya ke yang lain dengan eksositosis, memindahkan bagian-bagian penting dari membran (Gbr. 1.12.E), memberikan sel kesempatan, misalnya , membentuk hasil atau bergerak.
Penataan ulang serupa juga khas untuk sitoskeleton, terutama untuk mikrofilamen dan mikrotubulus (Gbr. 1.1). Mikrofilamen terutama terdiri dari Protein F-aktin yang mampu merakit menjadi bundel berserat sebagai hasil polimerisasi monomer dari sitosol. Bundel terpolarisasi, yaitu mereka sering tumbuh dari satu ujung saja, mengumpulkan molekul aktin baru, sedangkan ujung lainnya lembam atau pembongkaran terjadi di sini. Karena pertumbuhan terpolarisasi ini, mikrofilamen bergerak secara efisien dan struktur jaringannya dapat berubah. Transisi aktin dari keadaan terdepolimerisasi (sol) ke keadaan terorganisir (gel) dapat terjadi dengan sangat cepat di bawah pengaruh protein lain atau perubahan konsentrasi ion (lihat di bawah). Ada juga protein yang menyebabkan filamen aktin terurai menjadi fragmen pendek. Pertumbuhan tipis dari banyak sel - filopodia - mengandung bundel pusat aktin (Gbr. 1.1), dan berbagai gerakan filopodia mungkin disebabkan oleh transisi aktin: polimerisasi - depolimerisasi.
mikrotubulusjuga sering mengalami gerakan serupa. Mekanisme gerakan ini serupa - polimerisasi tubulin dari sitosol sedemikian rupa sehingga salah satu ujung mikrotubulus tumbuh, sementara yang lain tidak berubah, atau pembongkaran terjadi di sana. Dengan demikian, mikrotubulus, dengan penambahan atau penghilangan bahan yang tepat, dapat bergerak melalui sitosol.
Gerakan aktif sitoskeleton . Perubahan struktur sitoskeletal dapat terjadi sebagai akibat dari gerakan aktif dan penataan ulang yang dijelaskan di atas. Dalam banyak kasus, pergerakan mikrotubulus dan filamen aktin didorong oleh protein kontraktil yang mengikat filamen atau tubulus dan dapat memindahkannya relatif satu sama lain. tupai miosin dan dynein hadir dalam sitosol semua sel dalam konsentrasi yang relatif tinggi; mereka adalah elemen yang mengubah energi menjadi gerakan dalam sel khusus (otot) dan organel (silia). Dalam sel otot, miosin membentuk filamen tebal yang berorientasi sejajar dengan filamen aktin. Molekul miosin, dengan "kepalanya", menempel pada filamen aktin dan, menggunakan energi ATP, memindahkan miosin sepanjang molekul aktin. Myosin kemudian terlepas dari aktin. Himpunan banyak siklus koneksi-putus seperti itu mengarah ke makroskopik kontraksi serat otot(Bab 4). Dynein memainkan peran serupa dalam pergerakan mikrotubulus selama operasi silia (Gbr. 1.1). Dalam sitoplasma sel yang tidak terspesialisasi, miosin dan dynein tidak membentuk serat biasa, tetapi dalam banyak kasus kelompok molekul kecil. Bahkan dalam bentuk agregat kecil seperti itu, mereka mampu menggerakkan filamen aktin atau mikrotubulus. Beras. 1.13 mengilustrasikan proses ini ketika molekul miosin yang terpolarisasi berlawanan juga melekat pada dua filamen aktin yang terpolarisasi dalam arah yang berlawanan. Gugus kepala miosin menekuk ke arah ekor molekul, mengonsumsi ATP, dan kedua filamen aktin bergeser ke arah yang berlawanan, setelah itu miosin terlepas darinya. Pergerakan semacam ini, di mana energi ATP diubah menjadi kerja mekanis, dapat mengubah bentuk sitoskeleton dan, akibatnya, sel, serta menyediakan transportasi organel yang terkait dengan sitoskeleton.
Proses transpor intraseluler paling jelas ditunjukkan pada akson sel saraf. transportasi akson dibahas di sini secara rinci untuk menggambarkan peristiwa yang mungkin terjadi dengan cara yang sama di sebagian besar sel. Sebuah akson yang hanya berdiameter beberapa mikron dapat memiliki panjang satu meter atau lebih, dan akan memakan waktu bertahun-tahun bagi protein untuk berpindah melalui difusi dari nukleus ke ujung distal akson. Telah lama diketahui bahwa ketika setiap bagian akson mengalami penyempitan, bagian proksimal akson mengembang. Sepertinya aliran sentrifugal tersumbat di akson. Seperti aliran-cepat transportasi akson dapat ditunjukkan oleh pergerakan penanda radioaktif, seperti dalam percobaan yang ditunjukkan pada Gambar. 1.14. Leusin berlabel radioaktif disuntikkan ke ganglion akar dorsal, dan kemudian, dari jam ke-2 hingga ke-10, radioaktivitas diukur pada saraf siatik pada jarak 166 mm dari badan neuron. Selama 10 jam, puncak radioaktivitas di tempat suntikan tidak berubah secara signifikan. Namun gelombang radioaktivitas merambat di sepanjang akson dengan kecepatan konstan sekitar 34 mm per 2 jam, atau 410 mm/hari. Telah ditunjukkan bahwa di semua neuron hewan homoiothermic, transpor akson cepat terjadi pada kecepatan yang sama, dan tidak ada perbedaan mencolok antara serat tipis, tidak bermielin dan akson paling tebal, serta antara serat motorik dan sensorik. Jenis penanda radioaktif juga tidak mempengaruhi kecepatan transpor aksonal cepat; Berbagai zat radioaktif dapat berfungsi sebagai penanda.

Beras. 1.13.Kompleks miosin non-otot dengan orientasi tertentu dapat mengikat filamen aktin dengan polaritas yang berbeda dan, menggunakan energi ATP, menggesernya relatif satu sama lain.
molekul, seperti berbagai asam amino, yang dimasukkan ke dalam protein tubuh neuron. Jika kita menganalisis bagian perifer saraf untuk menentukan sifat pembawa radioaktivitas yang diangkut di sini, maka pembawa tersebut ditemukan terutama dalam fraksi protein, tetapi juga dalam komposisi mediator dan asam amino bebas. Mengetahui bahwa sifat-sifat zat ini berbeda dan terutama ukuran molekulnya berbeda, kita dapat menjelaskan laju transpor yang konstan hanya dengan mekanisme transpor yang umum untuk mereka semua.
Dijelaskan di atas transportasi akson cepat adalah anterograde yaitu diarahkan menjauh dari badan sel. Telah ditunjukkan bahwa beberapa zat bergerak dari perifer ke badan sel dengan bantuan: transportasi mundur. Misalnya, asetilkolinesterase diangkut ke arah ini dengan kecepatan dua kali lebih rendah daripada kecepatan transpor aksonal cepat. Penanda yang sering digunakan dalam neuroanatomi—peroksidase lobak—juga bergerak mundur. Transportasi retrograde mungkin memainkan peran penting dalam regulasi sintesis protein dalam tubuh sel. Beberapa hari setelah transeksi akson, kromatolisis diamati di badan sel, yang menunjukkan pelanggaran sintesis protein. Waktu yang diperlukan untuk kromatolisis berkorelasi dengan durasi transpor retrograde dari tempat transeksi akson ke badan sel. Hasil seperti itu juga menunjukkan penjelasan untuk pelanggaran ini - transmisi dari pinggiran "zat sinyal" yang mengatur sintesis protein terganggu. Jelas, "kendaraan" utama yang digunakan untuk aksonal cepat
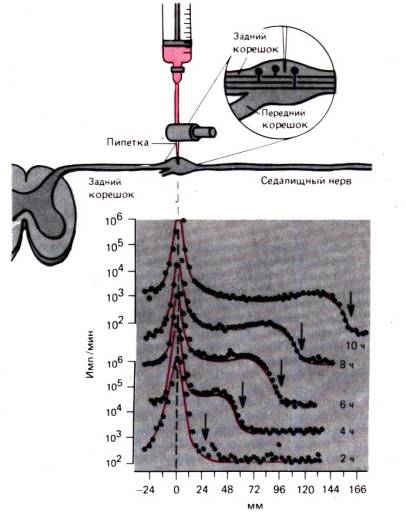
Beras. 1.14.Eksperimen yang mendemonstrasikan transpor akson cepat dalam serat sensorik saraf skiatik kucing. Leusin berlabel tritium disuntikkan ke ganglion akar dorsal dan radioaktivitas di ganglion dan serat sensorik diukur 2, 4, 6, 8, dan 10 jam setelah injeksi. (bagian bawah gambar). Oleh absis jarak dari ganglion ke bagian saraf siatik, tempat pengukuran dilakukan, ditunda. Pada sumbu y, hanya untuk kurva atas dan bawah, radioaktivitas (imp./min) diplot pada skala logaritmik. "Gelombang" radioaktivitas yang meningkat (panah) bergerak dengan kecepatan 410mm/hari (by )
transportasi adalah vesikel (vesikel) dan organel, seperti mitokondria yang mengandung zat yang akan diangkut. Pergerakan vesikel atau mitokondria terbesar dapat diamati menggunakan mikroskop in vivo . Partikel tersebut membuat gerakan pendek dan cepat dalam satu arah, berhenti, sering bergerak sedikit ke belakang atau ke samping, berhenti lagi, dan kemudian membuat lari ke arah utama. 410 mm/hari sesuai dengan kecepatan rata-rata anterograde sekitar 5 m/s; kecepatan setiap gerakan individu karenanya harus jauh lebih tinggi, dan jika kita memperhitungkan ukuran organel, filamen, dan mikrotubulus, maka gerakan ini benar-benar sangat cepat. Transportasi akson yang cepat membutuhkan konsentrasi ATP yang signifikan. Racun seperti kolkisin penghancur mikrotubulus juga memblok transpor aksonal yang cepat. Dari sini dapat disimpulkan bahwa dalam proses transportasi yang sedang kita pertimbangkan, vesikel dan organel bergerak di sepanjang mikrotubulus dan filamen aktin; gerakan ini disediakan oleh agregat kecil molekul dynein dan miosin yang bertindak seperti yang ditunjukkan pada gambar. 1.13, menggunakan energi ATP.
Transportasi akson cepat mungkin juga terlibat dalam proses patologis. Beberapa virus neurotropik (misalnya, virus herpes atau polio) menembus akson di perifer dan bergerak melalui transpor retrograde ke badan neuron, tempat virus tersebut berkembang biak dan memberikan efek toksiknya. Toksin tetanus, protein yang diproduksi oleh bakteri yang masuk ke dalam tubuh melalui lesi kulit, diambil oleh ujung saraf dan diangkut ke badan neuron, di mana ia menyebabkan kejang otot yang khas. Kasus efek toksik pada transportasi akson itu sendiri diketahui, misalnya, paparan pelarut industri akrilamida. Selain itu, diyakini bahwa patogenesis beri-beri beri-beri dan polineuropati alkoholik termasuk pelanggaran transportasi aksonal cepat.
Selain transportasi akson yang cepat di dalam sel, ada juga yang agak intens transportasi akson lambat. Tubulin bergerak di sepanjang akson dengan kecepatan sekitar 1 mm/hari, sementara aktin bergerak lebih cepat, hingga 5 mm/hari. Protein lain juga bermigrasi dengan komponen sitoskeleton ini; misalnya, enzim tampaknya terkait dengan aktin atau tubulin. Laju pergerakan tubulin dan aktin secara kasar konsisten dengan laju pertumbuhan yang ditemukan untuk mekanisme yang dijelaskan sebelumnya ketika molekul digabungkan ke ujung aktif mikrotubulus atau mikrofilamen. Oleh karena itu, mekanisme ini mungkin mendasari transportasi aksonal yang lambat. Laju transpor akson yang lambat juga kira-kira sesuai dengan laju pertumbuhan akson, yang, tampaknya, menunjukkan keterbatasan yang dipaksakan oleh struktur sitoskeleton pada proses kedua.
Sebagai penutup bagian ini, harus ditekankan bahwa sel sama sekali bukan struktur statis, seperti yang tampak, misalnya, dalam foto mikroskopis elektron. membran plasma dan terutama organel berada dalam gerakan cepat yang konstan dan restrukturisasi konstan; itulah satu-satunya alasan mereka dapat berfungsi. Selanjutnya, ini bukan ruang sederhana di mana reaksi kimia berlangsung, tetapi konglomerat membran dan serat yang sangat terorganisir, di mana reaksi berlangsung dalam urutan yang terorganisir secara optimal.
Pemeliharaan sel individu sebagai unit fungsional sebagian besar diatur oleh nukleus; studi tentang mekanisme pengaturan tersebut adalah subjek biologi sel dan biokimia. Pada saat yang sama, sel-sel harus memodifikasi fungsinya sesuai dengan kondisi lingkungan dan kebutuhan sel-sel tubuh lainnya, yaitu, mereka berfungsi sebagai objek regulasi fungsional. Di bawah ini kami secara singkat mempertimbangkan bagaimana pengaruh pengaturan ini bekerja pada membran plasma dan bagaimana mereka mencapai organel intraseluler.
Efek regulasi pada membran sel
Potensial membran . Dalam banyak kasus, pengaturan fungsi seluler dilakukan dengan mengubah potensial membran. Perubahan potensial lokal dimungkinkan ketika: 1) arus dari area sel tetangga atau dihasilkan oleh sel lain mengalir melalui membran; 2) konsentrasi ion ekstraseluler berubah (seringkali [K + ] keluar ); 3) saluran ion membran terbuka. Perubahan potensial membran dapat mempengaruhi konformasi protein membran, menyebabkan, khususnya, saluran untuk membuka atau menutup. Seperti dijelaskan di atas, fungsi beberapa pompa membran bergantung pada potensial membran. Sel saraf terspesialisasi untuk merasakan perubahan potensial membran sebagai informasi yang harus diproses dan ditransmisikan (lihat Bab 2).
zat pengatur ekstraseluler . Mekanisme pengaturan terpenting yang melibatkan zat ekstraseluler adalah interaksinya dengan reseptor spesifik pada membran plasma atau di dalam sel. Zat-zat ini termasuk mediator sinaptik yang mengirimkan informasi antara sel-sel saraf, agen lokal dan zat yang beredar dalam darah dan mencapai semua sel tubuh, seperti hormon dan antigen. neurotransmiter sinaptik adalah molekul kecil yang dilepaskan dari ujung saraf di sinaps;
ketika mereka mencapai membran plasma sel postsinaptik tetangga, mereka memicu sinyal listrik atau mekanisme pengaturan lainnya. Masalah ini dibahas secara rinci dalam Bab. 3.
Agen kimia lokal sering disekresikan oleh sel-sel khusus. Mereka berdifusi secara bebas di ruang ekstraseluler, tetapi aksinya terbatas pada sekelompok kecil sel karena penghancuran cepat zat-zat ini, baik secara spontan atau oleh aksi enzim. Salah satu contoh pelepasan agen tersebut adalah rilis histamin sel mast pada cedera atau respon imun. Histamin menyebabkan relaksasi sel otot polos pembuluh darah, meningkatkan permeabilitas endotel pembuluh darah, dan merangsang ujung saraf sensorik yang memediasi sensasi gatal. Agen kimia lokal lainnya disekresikan oleh banyak sel lain. Agen lokal yang khas adalah prostaglandin, merupakan kelompok sekitar 20 turunan asam lemak. Mereka dilepaskan terus menerus dari sel yang tersebar luas, tetapi hanya bekerja secara lokal, karena mereka dengan cepat dihancurkan oleh fosfolipase membran. Berbagai prostaglandin memiliki spektrum aksi yang luas: mereka dapat memicu kontraksi sel otot polos, menyebabkan agregasi trombosit (trombosit), atau menghambat perkembangan korpus luteum di ovarium.
Agen lokal lainnya melayani faktor pertumbuhan. Faktor pertumbuhan saraf (NGF) yang paling terkenal untuk neuron simpatis, yang diperlukan untuk pertumbuhan dan kelangsungan hidup neuron ini selama perkembangan in vivo atau dalam kultur sel. Jelas, sel target untuk kelas neuron ini mengeluarkan NGF dan dengan demikian memberikan persarafan yang benar. Saat membentuk organ, sel seringkali perlu "menemukan jalan" untuk menargetkan sel, yang dapat ditempatkan pada jarak yang cukup jauh. Oleh karena itu, harus ada banyak faktor pertumbuhan khusus seperti NGF.
Hormon dan antigen dibawa oleh darah ke semua sel. Antigen menimbulkan respon imun dari sel yang membawa antibodi spesifik. Namun, antigen, sebagai aturan, adalah zat asing yang tidak terbentuk dalam organisme yang bereaksi (untuk lebih jelasnya, lihat Bab 18). Beberapa hormon, seperti insulin atau tiroksin, mempengaruhi berbagai jenis sel, sementara yang lain, seperti hormon seks, hanya mempengaruhi jenis sel tertentu. Hormon adalah peptida, yang aksinya dipicu oleh ikatannya dengan reseptor pada membran sel, atau steroid dan tiroksin, yang berdifusi melalui membran lipid dan berikatan dengan reseptor intraseluler. Hormon steroid mengikat kromatin inti, menghasilkan transkripsi gen tertentu. Protein yang dihasilkan sebagai akibatnya menyebabkan perubahan fungsi seluler, yang merupakan efek spesifik dari hormon. Isu-isu yang berkaitan dengan pelepasan dan kerja hormon dibahas secara rinci dalam Bab. 17.
Komunikasi intraseluler yang melibatkan utusan kedua
Fungsi pengaturan yang dijelaskan di atas termasuk efek pada membran sel. Informasi yang diterima oleh membran sel harus sering menyebabkan reaksi di organel dan dibawa ke mereka oleh berbagai zat yang dikenal sebagai utusan kedua (sebagai lawan yang pertama, datang ke sel dari sumber eksternal). Studi tentang mediator kedua berkembang pesat, dan tidak ada jaminan bahwa tingkat pemahaman masalah saat ini akan cukup lengkap. Di sini kita akan membahas tiga mediator yang telah dipelajari dengan baik: Ca 2+ , cAMP dan inositol triphosphate.
Kalsium.Mediator intraseluler yang paling sederhana adalah ion Ca2+. Konsentrasi bebasnya dalam sel istirahat sangat rendah dan berjumlah 10 -8 -10 -7 mol/l. Ia dapat masuk ke dalam sel melalui saluran membran tertentu ketika mereka terbuka, misalnya, ketika potensial membran berubah (lihat Bab 2). Peningkatan Ca2+ yang dihasilkan memicu reaksi penting dalam sel, seperti kontraksi miofibril, yang merupakan dasar kontraksi otot (lihat Bab 4), atau pelepasan vesikel yang mengandung neurotransmiter dari ujung saraf (lihat Bab 3) . Kedua reaksi memerlukan konsentrasi Ca 2+ sekitar 10–5 mol/l. Ca 2+ , yang memiliki efek pengaturan, juga dapat dilepaskan dari depot intraseluler, seperti retikulum endoplasma. Pelepasan Ca 2+ dari depot memerlukan partisipasi perantara lain (lihat, misalnya, Gambar 1.16).
Siklik adenosin monofosfat, cAMP. Baru-baru ini, telah terbukti bahwa siklik adenosin monofosfat (cAMP), turunan dari sumber energi utama dalam tubuh, ATP, merupakan utusan kedua yang penting. Rantai reaksi kompleks yang ditunjukkan pada gambar. 1.15, dimulai dari reseptor Rp pada permukaan luar membran plasma, yang dapat berfungsi sebagai tempat pengikatan spesifik untuk berbagai mediator dan hormon. Setelah mengikat molekul "stimulasi" tertentu Rp mengubah konformasinya; perubahan ini mempengaruhi protein Gs pada permukaan bagian dalam membran sedemikian rupa sehingga menjadi mungkin untuk mengaktifkan yang terakhir oleh guanosin trifosfat intraseluler (GTP). protein aktif Gs , pada gilirannya, merangsang enzim pada permukaan bagian dalam membran, adenilat siklase (AC), yang mengkatalisis pembentukan cAMP dari ATP. cAMP yang larut dalam air dan merupakan mediator yang mentransmisikan efek

Beras. 1.15.Sebuah rantai reaksi yang melibatkan cAMP mediator intraseluler (siklik adenosin monofosfat). Sinyal eksternal rangsang atau penghambatan mengaktifkan reseptor membran R s atau Ri . Reseptor ini mengatur proses pengikatan G -protein dengan GTP intraseluler (guanosin trifosfat), sehingga merangsang atau menghambat adenilat siklase (AC) intraseluler. Enzim penguat AC mengubah adenosin trifosfat (ATP) menjadi cAMP, yang kemudian dipecah menjadi AMP dengan bantuan fosfodiester. cAMP bebas berdifusi ke dalam sel dan mengaktifkan adenilat kinase (A-kinase), melepaskan subunit katalitik C, yang mengkatalisis fosforilasi protein intraseluler, mis. membentuk efek akhir dari stimulus ekstraseluler. Skema ini juga menunjukkan obat farmakologis dan racun yang memicu (+) atau menghambat (-) beberapa reaksi (tetapi dengan perubahan)
stimulasi reseptor ekstraseluler Rp terhadap struktur internal sel.
Sejalan dengan rantai reaksi stimulasi yang melibatkan Rp kemungkinan pengikatan mediator penghambat dan hormon ke reseptor yang sesuai R i yang lagi-lagi melalui protein yang diaktifkan GTP G , menghambat AC dan dengan demikian produksi cAMP. Difusi ke dalam sel, cAMP bereaksi dengan adenilat kinase (A-kinase); Ini melepaskan subunit C, yang mengkatalisis fosforilasi protein. Fosforilasi ini mengubah protein menjadi bentuk aktifnya, dan sekarang mereka dapat mengerahkan tindakan pengaturan spesifiknya (misalnya, menyebabkan degradasi glikogen). Sistem regulasi yang kompleks ini sangat efisien, karena hasil akhirnya adalah fosforilasi banyak protein, yaitu, sinyal regulasi melewati rantai dengan faktor amplifikasi yang besar. Mediator eksternal yang berikatan dengan reseptor R s dan R i spesifik untuk masing-masing sangat beragam. Adrenalin, bermain-main dengan R s atau R i berpartisipasi dalam regulasi metabolisme lipid dan glikogen, serta dalam peningkatan kontraksi otot jantung dan reaksi lainnya (lihat Bab 19). Hormon perangsang kelenjar gondok, mengaktifkan Rp , merangsang sekresi hormon tiroksin oleh kelenjar tiroid, dan prostaglandin I menghambat agregasi trombosit. Efek penghambatan, termasuk adrenalin, dimediasi melalui R i diekspresikan dalam memperlambat lipolisis. Lewat sini, sistem cAMP adalah sistem pengaturan intraseluler multifungsi, yang dapat dikontrol secara tepat oleh agen pensinyalan stimulator dan penghambat ekstraseluler.
Inositol fosfat "JIKA" H ". Sistem intraseluler dari mediator kedua, inositol fosfat, baru-baru ini ditemukan (Gbr. 1.16). Dalam hal ini, tidak ada jalur penghambatan, tetapi ada kesamaan dengan sistem cAMP, di mana efek stimulasi reseptor R ditransfer ke protein G yang diaktifkan GTP di permukaan bagian dalam membran. Pada tahap berikutnya, fosfatidilinositol (PI) lipid membran biasa, yang sebelumnya menerima dua gugus fosfat tambahan, diubah menjadi PI-difosfat (FIF 2), yang dipecah oleh fosfodiesterase teraktivasi (PDE) menjadi inositol trifosfat(IFz) dan lipid diasilgliserol(DAG). Inositol trifosfat adalah pembawa pesan kedua yang larut dalam air yang berdifusi ke dalam sitosol. Bekerja terutama dengan melepaskan Ca2+ dari retikulum endoplasma. Ca 2+ pada gilirannya bertindak sebagai perantara seperti yang dijelaskan di atas; misalnya, ia mengaktifkan fosfokinase yang bergantung pada Ca2+, yang memfosforilasi enzim. Subunit lipid DAG (Gbr. 1.16) juga membawa sinyal, berdifusi dalam fase lipid membran plasma ke C-kinase yang terletak di permukaan dalamnya, yang diaktifkan dengan partisipasi fosfatidilserin sebagai kofaktor. Kemudian C-kinase memicu fosforilasi protein, mengubahnya menjadi bentuk aktif.
Sistem intraseluler dari interferon 3 perantara kedua juga dapat dikendalikan oleh berbagai mediator dan hormon eksternal, termasuk asetilkolin, serotonin, vasopresin, dan hormon perangsang tiroid; seperti sistem cAMP, ini ditandai dengan berbagai efek intraseluler. Ada kemungkinan bahwa sistem ini juga diaktifkan oleh cahaya di reseptor visual mata dan memainkan peran sentral dalam fototransduksi (lihat Bab 11). Untuk pertama kalinya dalam perkembangan individu suatu organisme, reseptor sistem IGF diaktifkan oleh sperma, akibatnya IGF mengambil bagian dalam reaksi regulasi yang menyertai pembuahan sel telur.
Sistem cAMP dan IFz-DAG sangat efektif penambah biologis. Mereka

Beras. 1.16.Sebuah rantai reaksi yang melibatkan interferon perantara intraseluler (inositol trifosfat). Seperti dalam sistem cAMP, sinyal ekstraseluler dimediasi melalui proteinG, yang dalam hal ini mengaktifkan fosfodiesterase (PDE). Enzim ini memecah phosphatidylinosine diphosphate (FIF) 2 ) di membran plasma sebelum IF H dan diasilgliserol (DAG); JIKA H berdifusi ke dalam sitoplasma. Di sini dia menyebabkan pelepasan Ca 2+ dari retikulum endoplasma; peningkatan konsentrasi Ca 2+ dalam sitoplasma ([Ca 2+ ] saya ) mengaktifkan protein kinase yang memfosforilasi dan oleh karena itu mengaktifkan enzim. Produk lain, DAG, tetap berada di membran dan mengaktifkan protein kinase C (kofaktor-fosfatidilserin, PS). Protein kinase C juga memfosforilasi enzim yang memediasi tindakan spesifik yang terkait dengan stimulasi reseptor eksternal. R . Cabang-cabang rantai reaksi yang melibatkan IF H dan DAG dapat diaktifkan secara independen oleh ionomycin dan phorbol ester, masing-masing (dimodifikasi)
mengubah reaksi antara mediator dan reseptor membran luar menjadi fosforilasi banyak protein intraseluler, yang kemudian dapat mempengaruhi berbagai fungsi sel. Salah satu aspek penting dari masalah ini adalah, sejauh yang kita ketahui saat ini, hanya ada dua sistem regulasi yang terkait erat dari jenis ini, yang digunakan oleh banyak mediator eksternal untuk mengatur berbagai proses intraseluler. Pada saat yang sama, sistem pengaturan ini, termasuk Ca 2+ , berinteraksi erat satu sama lain, yang memungkinkan mereka untuk melakukan pengaturan fungsi seluler yang baik.
Tutorial dan panduan
1. Alberts DI DALAM., Bray D., Lewis J., Raff M., Roberts KE., Watson J.D.Biologi Molekuler Sel, New York dan London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biologi. Berlin, Heidelberg, New York, Springer, 1983.
3. bukit DI DALAM. Saluran ionik dari membran yang dapat dieksitasi. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biofisika. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann KE., Mahler H.Biokimia. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E.R., Schwartz-J.H.,(eds.). Prinsip-prinsip ilmu saraf, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., SchmidtW.Anatomi des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Artikel dan ulasan asli
8. Berridge M.J. Dasar molekuler komunikasi di dalam sel, Sci. Amer 253 124 134 (1985).
9. Berridge M. J., Irvine R. F. Inositol triphosphate, utusan kedua baru dalam transduksi sinyal seluler. Alam, 312, 315 321 (1984).
10. Bretscher M.S. Molekul membran sel, Sci. Amer., 253, 124–134 (1985).
11. Daut J. Sel hidup sebagai mesin transduksi energi. Model minimal metabolisme miokard, Biochem. dan Biofis. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz DI DALAM. Pengaruh ion natrium pada aktivitas listrik akson raksasa cumi-cumi. J Fisiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Transpor aktif kation dalam akson raksasa dari Warna coklat tua dan loligo, J Fisiol. (Land.), 128, 28–42 (1955).
14. lebih lama P Saluran ionik dengan substat konformasi, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Senilai P.M. Transpor eksoplasma dalam sistem normal dan patologis. Dalam: Fisiologi dan Patologi Akson, S.G. tukang lilin, Ed. New York, Raven Press, 1978.
Kami mengundang Anda untuk membiasakan diri dengan materi dan.
: membran selulosa, membran, sitoplasma dengan organel, nukleus, vakuola dengan getah sel.Kehadiran plastida adalah fitur utama dari sel tumbuhan.
Fungsi dinding sel- menentukan bentuk sel, melindungi terhadap faktor lingkungan.
membran plasma- film tipis, terdiri dari molekul lipid dan protein yang berinteraksi, membatasi konten internal dari lingkungan eksternal, menyediakan transportasi air, mineral dan zat organik ke dalam sel melalui osmosis dan transfer aktif, dan juga menghilangkan produk limbah.
sitoplasma- lingkungan semi-cair internal sel, di mana nukleus dan organel berada, menyediakan koneksi di antara mereka, berpartisipasi dalam proses utama kehidupan.
Retikulum endoplasma- jaringan saluran percabangan di sitoplasma. Ini terlibat dalam sintesis protein, lipid dan karbohidrat, dalam pengangkutan zat. Ribosom - badan yang terletak di EPS atau di sitoplasma, terdiri dari RNA dan protein, terlibat dalam sintesis protein. EPS dan ribosom adalah alat tunggal untuk sintesis dan transportasi protein.
Mitokondria-organel dipisahkan dari sitoplasma oleh dua membran. Zat organik dioksidasi di dalamnya dan molekul ATP disintesis dengan partisipasi enzim. Peningkatan permukaan membran bagian dalam tempat enzim berada karena krista. ATP adalah zat organik yang kaya energi.
plastida(kloroplas, leukoplas, kromoplas), kandungannya dalam sel adalah ciri utama organisme tumbuhan. Kloroplas adalah plastida yang mengandung pigmen hijau klorofil, yang menyerap energi cahaya dan menggunakannya untuk mensintesis zat organik dari karbon dioksida dan air. Pembatasan kloroplas dari sitoplasma oleh dua membran, banyak hasil - grana pada membran bagian dalam, di mana molekul klorofil dan enzim berada.
Kompleks Golgi- sistem rongga yang dibatasi dari sitoplasma oleh membran. Akumulasi protein, lemak dan karbohidrat di dalamnya. Implementasi sintesis lemak dan karbohidrat pada membran.
Lisosom- tubuh dipisahkan dari sitoplasma oleh membran tunggal. Enzim yang terkandung di dalamnya mempercepat reaksi pemecahan molekul kompleks menjadi yang sederhana: protein menjadi asam amino, karbohidrat kompleks menjadi sederhana, lipid menjadi gliserol dan asam lemak, dan juga menghancurkan bagian sel yang mati, seluruh sel.
Vakuola- rongga di sitoplasma diisi dengan getah sel, tempat akumulasi nutrisi cadangan, zat berbahaya; mereka mengatur kadar air dalam sel.
Inti- bagian utama sel, ditutupi di luar dengan dua membran, ditembus oleh selubung inti pori-pori. Zat memasuki inti dan dikeluarkan darinya melalui pori-pori. Kromosom adalah pembawa informasi herediter tentang karakteristik suatu organisme, struktur utama nukleus, yang masing-masing terdiri dari satu molekul DNA dalam kombinasi dengan protein. Nukleus merupakan tempat sintesis DNA, i-RNA, r-RNA.

Adanya membran luar, sitoplasma dengan organel, nukleus dengan kromosom.
Membran luar atau plasma- membatasi isi sel dari lingkungan (sel lain, zat antar sel), terdiri dari molekul lipid dan protein, menyediakan komunikasi antar sel, pengangkutan zat ke dalam sel (pinositosis, fagositosis) dan keluar sel.
sitoplasma- lingkungan semi-cair internal sel, yang menyediakan komunikasi antara nukleus dan organel yang terletak di dalamnya. Proses utama aktivitas vital terjadi di sitoplasma.
Organel sel:
1) retikulum endoplasma (RE)- sistem tubulus bercabang, yang terlibat dalam sintesis protein, lipid dan karbohidrat, dalam pengangkutan zat dalam sel;
2) ribosom- badan yang mengandung rRNA terletak di RE dan di sitoplasma, dan terlibat dalam sintesis protein. EPS dan ribosom adalah alat tunggal untuk sintesis dan transportasi protein;
3) mitokondria- "pembangkit tenaga" sel, dipisahkan dari sitoplasma oleh dua membran. Bagian dalam membentuk krista (lipatan) yang meningkatkan permukaannya. Enzim pada krista mempercepat reaksi oksidasi zat organik dan sintesis molekul ATP yang kaya energi;
4) kompleks golgi- sekelompok rongga yang dibatasi oleh membran dari sitoplasma, diisi dengan protein, lemak, dan karbohidrat, yang digunakan dalam proses kehidupan atau dikeluarkan dari sel. Membran kompleks melakukan sintesis lemak dan karbohidrat;
5) lisosom- tubuh yang diisi dengan enzim mempercepat reaksi pemecahan protein menjadi asam amino, lipid menjadi gliserol dan asam lemak, polisakarida menjadi monosakarida. Dalam lisosom, bagian sel yang mati, seluruh sel dan sel dihancurkan.
Inklusi sel- Akumulasi nutrisi cadangan: protein, lemak dan karbohidrat.
Inti- bagian terpenting dari sel. Itu ditutupi dengan membran membran ganda dengan pori-pori di mana beberapa zat menembus ke dalam nukleus, sementara yang lain memasuki sitoplasma. Kromosom adalah struktur utama nukleus, pembawa informasi turun-temurun tentang karakteristik suatu organisme. Ini ditransmisikan dalam proses pembelahan sel induk ke sel anak, dan dengan sel germinal - ke organisme anak. Nukleus adalah tempat sintesis DNA, mRNA, rRNA.
Tugas:
Jelaskan mengapa organel disebut struktur khusus sel?
Menjawab: organel disebut struktur sel khusus, karena mereka melakukan fungsi yang ditentukan secara ketat, informasi herediter disimpan dalam nukleus, ATP disintesis dalam mitokondria, fotosintesis berlangsung di kloroplas, dll.
Jika Anda memiliki pertanyaan tentang sitologi, Anda dapat meminta bantuan dari
Tahap ketiga evolusi adalah penampilan sel.
Molekul protein dan asam nukleat (DNA dan RNA) membentuk sel biologis, unit terkecil kehidupan. Sel biologis adalah "blok bangunan" dari semua organisme hidup dan mengandung semua kode material perkembangan.
Untuk waktu yang lama, para ilmuwan menganggap struktur sel sangat sederhana. Kamus Ensiklopedis Soviet menafsirkan konsep sel sebagai berikut: "Sel adalah sistem kehidupan dasar, dasar dari struktur dan kehidupan semua hewan dan tumbuhan." Perlu dicatat bahwa istilah "dasar" sama sekali tidak berarti "sederhana". Sebaliknya, sel adalah ciptaan Tuhan yang fraktal, mencolok dalam kompleksitasnya dan, pada saat yang sama, dalam koherensi pekerjaan yang luar biasa. dari semua elemennya.
Ketika kami berhasil melihat ke dalam dengan bantuan mikroskop elektron, ternyata perangkat sel paling sederhana sama kompleks dan tidak dapat dipahami seperti Semesta itu sendiri. Hari ini telah ditetapkan bahwa "Sel adalah materi khusus Semesta, materi khusus Kosmos." Satu sel tunggal berisi informasi yang hanya dapat dimasukkan ke dalam beberapa puluh ribu volume Ensiklopedia Besar Soviet. Itu. sel, antara lain, adalah "bioreservoir" informasi yang sangat besar.
Penulis teori evolusi molekuler modern, Manfred Eigen, menulis: “Agar molekul protein terbentuk secara kebetulan, alam harus melakukan sekitar 10130 percobaan dan menghabiskan jumlah molekul yang cukup untuk 1027 percobaan. Semesta. Jika protein dibangun dengan cerdas, yaitu bahwa validitas setiap gerakan dapat diperiksa oleh beberapa mekanisme seleksi, hanya dibutuhkan sekitar 2000 upaya. Kami sampai pada kesimpulan paradoks: program untuk membangun "sel hidup primitif" dikodekan di suatu tempat pada tingkat partikel dasar ".
Dan bagaimana bisa sebaliknya. Setiap sel, yang memiliki DNA, diberkahi dengan kesadaran, sadar akan dirinya sendiri dan sel-sel lain, dan berhubungan dengan Semesta, pada kenyataannya, merupakan bagian darinya. Dan meskipun jumlah dan variasi sel dalam tubuh manusia luar biasa (sekitar 70 triliun), mereka semua serupa, sama seperti semua proses yang terjadi dalam sel serupa. Dalam kata-kata ilmuwan Jerman Roland Glaser, desain sel biologis "dipikirkan dengan sangat baik." Siapa yang dipikirkan dengan baik?
Jawabannya sederhana: protein, asam nukleat, sel hidup dan semua sistem biologis adalah produk dari aktivitas kreatif Pencipta intelektual.
Yang menarik: pada tingkat atom, tidak ada perbedaan antara komposisi kimia dunia organik dan anorganik. Dengan kata lain, pada tingkat atom, sel diciptakan dari unsur-unsur yang sama seperti alam mati. Perbedaan ditemukan pada tingkat molekuler. Di dalam tubuh makhluk hidup, selain zat anorganik dan air, juga terdapat protein, karbohidrat, lemak, asam nukleat, enzim ATP sintase, dan senyawa organik berbobot molekul rendah lainnya.
Sampai saat ini, sel telah benar-benar dibongkar menjadi atom untuk tujuan studi. Namun, tidak mungkin membuat setidaknya satu sel hidup, karena membuat sel berarti membuat partikel Alam Semesta yang hidup. Akademisi V.P. Kaznacheev percaya bahwa "sel adalah organisme kosmoplanet ... Sel manusia adalah sistem tertentu dari biocollider torsi halus. Dalam biocollider ini, proses yang tidak kita ketahui terjadi, materialisasi bentuk aliran kosmik terjadi, transformasi kosmiknya dan karena ini partikel terwujud".
Air.
Hampir 80% dari massa sel adalah air. Menurut Doktor Biologi S. Zenin, air, karena struktur clusternya, merupakan matriks informasi untuk mengelola proses biokimia. Selain itu, airlah yang merupakan "target" utama yang berinteraksi dengan osilasi frekuensi suara. Keteraturan air seluler sangat tinggi (mendekati keteraturan kristal) sehingga disebut kristal cair.
tupai.
Protein memainkan peran penting dalam kehidupan biologis. Sel mengandung beberapa ribu protein yang unik untuk jenis sel ini (dengan pengecualian sel punca). Kemampuan untuk mensintesis proteinnya sendiri diwariskan dari sel ke sel dan bertahan sepanjang hidup. Selama kehidupan sel, protein secara bertahap mengubah strukturnya, fungsinya terganggu. Protein bekas ini dikeluarkan dari sel dan diganti dengan yang baru, berkat aktivitas vital sel yang dipertahankan.
Pertama-tama, kami mencatat fungsi bangunan protein, karena mereka adalah bahan bangunan yang membentuk membran sel dan organel seluler, dinding pembuluh darah, tendon, tulang rawan, dll.
Fungsi sinyal protein sangat menarik. Ternyata protein mampu berfungsi sebagai zat pemberi sinyal, mentransmisikan sinyal antar jaringan, sel atau organisme. Fungsi pensinyalan dilakukan oleh protein hormon. Sel dapat berkomunikasi satu sama lain dari jarak jauh menggunakan protein pensinyalan yang ditransmisikan melalui zat antar sel.
Protein juga memiliki fungsi motorik. Semua jenis gerakan yang mampu dilakukan sel, seperti kontraksi otot, dilakukan oleh protein kontraktil khusus. Protein juga melakukan fungsi transportasi. Mereka mampu menempelkan berbagai zat dan memindahkannya dari satu tempat di dalam sel ke tempat lain. Misalnya, protein darah hemoglobin mengikat oksigen dan membawanya ke seluruh jaringan dan organ tubuh. Selain itu, protein juga memiliki fungsi protektif. Ketika protein atau sel asing dimasukkan ke dalam tubuh, protein khusus diproduksi di dalamnya yang mengikat dan menetralkan sel dan zat asing. Dan akhirnya, fungsi energi protein adalah bahwa dengan pemecahan lengkap 1 g protein, energi dilepaskan dalam jumlah 17,6 kJ.
Struktur sel.
Sel terdiri dari tiga bagian yang saling terkait: membran, sitoplasma dan nukleus, dan struktur dan fungsi nukleus dalam periode kehidupan sel yang berbeda berbeda. Untuk kehidupan sel mencakup dua periode: pembelahan, sebagai akibatnya dua sel anak terbentuk, dan periode antara pembelahan, yang disebut interfase.
Membran sel berinteraksi langsung dengan lingkungan luar dan berinteraksi dengan sel tetangga. Ini terdiri dari lapisan luar dan membran plasma yang terletak di bawahnya. Lapisan permukaan sel hewan disebut glikokalis. Ini menghubungkan sel dengan lingkungan eksternal dan dengan semua zat di sekitarnya. Ketebalannya kurang dari 1 mikron.
Struktur sel
Membran sel merupakan bagian yang sangat penting dari sel. Ini menyatukan semua komponen seluler dan membatasi lingkungan eksternal dan internal.
Ada pertukaran zat yang konstan antara sel dan lingkungan eksternal. Air, berbagai garam dalam bentuk ion individu, molekul anorganik dan organik memasuki sel dari lingkungan eksternal. Produk metabolisme, serta zat yang disintesis dalam sel: protein, karbohidrat, hormon, yang diproduksi dalam sel berbagai kelenjar, diekskresikan ke lingkungan eksternal melalui membran dari sel. Pengangkutan zat adalah salah satu fungsi utama membran plasma.
sitoplasma- media semi-cair internal di mana proses metabolisme utama berlangsung. Studi terbaru menunjukkan bahwa sitoplasma bukanlah sejenis larutan, komponen-komponen yang berinteraksi satu sama lain dalam tumbukan acak. Ini dapat dibandingkan dengan jeli, yang mulai "bergetar" sebagai respons terhadap pengaruh eksternal. Ini adalah bagaimana sitoplasma merasakan dan mengirimkan informasi.
Nukleus dan berbagai organel terletak di sitoplasma, yang disatukan olehnya menjadi satu kesatuan, yang memastikan interaksi mereka dan aktivitas sel sebagai sistem integral tunggal. Nukleus terletak di bagian tengah sitoplasma. Seluruh zona bagian dalam sitoplasma diisi dengan retikulum endoplasma, yang merupakan organoid seluler: sistem tubulus, vesikel, dan "tangki" yang dibatasi oleh membran. Retikulum endoplasma terlibat dalam proses metabolisme, menyediakan pengangkutan zat dari lingkungan ke sitoplasma dan antara struktur intraseluler individu, tetapi fungsi utamanya adalah partisipasi dalam sintesis protein, yang dilakukan di ribosom. - tubuh kecil mikroskopis berbentuk bulat dengan diameter 15-20 nm. Protein yang disintesis pertama kali terakumulasi dalam saluran dan rongga retikulum endoplasma dan kemudian diangkut ke organel dan situs sel di mana mereka dikonsumsi.
Selain protein, sitoplasma juga mengandung mitokondria, badan kecil berukuran 0,2-7 mikron, yang disebut "pembangkit tenaga" sel. Reaksi redoks terjadi di mitokondria, menyediakan sel dengan energi. Jumlah mitokondria dalam satu sel bervariasi dari beberapa hingga beberapa ribu.
Inti- bagian vital sel, mengontrol sintesis protein dan melalui mereka semua proses fisiologis dalam sel. Dalam nukleus sel yang tidak membelah, membran nukleus, jus nukleus, nukleolus, dan kromosom dibedakan. Melalui selubung nukleus, terjadi pertukaran zat yang terus menerus antara nukleus dan sitoplasma. Di bawah amplop nuklir - jus nuklir (zat semi-cair), yang mengandung nukleolus dan kromosom. Nukleolus adalah tubuh bulat padat, yang dimensinya dapat sangat bervariasi, dari 1 hingga 10 mikron dan lebih banyak lagi. Ini terutama terdiri dari ribonukleoprotein; berpartisipasi dalam pembentukan ribosom. Biasanya ada 1-3 nukleolus di dalam sel, kadang sampai beberapa ratus. Nukleolus terdiri dari RNA dan protein.
Dengan munculnya sel, Kehidupan muncul di Bumi!
Bersambung...


