Omogućava odnos ćelije sa okolinom. Veza organizma sa okolinom
Veza organizma sa okolinom, sa fizičko-hemijske tačke gledišta, je otvoren sistem, odnosno sistem u kome se odvijaju biohemijski procesi. Početne tvari dolaze iz okoline, a tvari koje se također kontinuirano formiraju se izvlače. Ravnoteža između brzine i koncentracije produkata višesmjernih reakcija u tijelu je uvjetna, imaginarna, jer unos i uklanjanje tvari ne prestaju. Kontinuirana povezanost sa okolinom i omogućava nam da posmatramo živi organizam kao otvoreni sistem.
Sunce je izvor energije za sve žive ćelije. Biljne ćelije hvataju energiju sunčeve svetlosti uz pomoć hlorofila, koristeći je za reakcije asimilacije tokom fotosinteze. Ćelije životinja, gljiva, bakterija koriste sunčevu energiju indirektno, kada cijepaju organske tvari koje sintetizira zemaljska biljka.
Dio hranljivih sastojaka ćelije se razgrađuje u procesu ćelijskog disanja, čime se snabdeva energijom neophodnom za različite vrste ćelijske aktivnosti. Ovaj proces se odvija u organelama zvanim mitohondrije. Mitohondrija se sastoji od dvije membrane: vanjske, koja odvaja organelu od citoplazme, i unutrašnje, koja formira brojne nabore. Glavni proizvod disanja je ATP. Napušta mitohondrije i koristi se kao izvor energije za mnoge hemijske reakcije u citoplazmi i ćelijskoj membrani. Ako je kisik potreban za provedbu ćelijskog disanja, tada se disanje naziva aerobno disanje, ali ako se reakcije odvijaju u nedostatku kisika, onda se govori o anaerobnom disanju.
Za bilo koju vrstu rada u ćeliji, energija se koristi u jednom obliku - u obliku energije iz fosfatnih veza ATP-a. ATP je vrlo mobilno jedinjenje. Formiranje ATP-a događa se na unutrašnjoj membrani mitohondrija. ATP se sintetiše u svim ćelijama tokom disanja zahvaljujući energiji oksidacije ugljenih hidrata, masti i drugih organskih materija. U ćelijama zelenih biljaka, glavna količina ATP-a se sintetizira u hloroplastima zahvaljujući sunčevoj energiji. U njima se tokom fotosinteze proizvodi višestruko više ATP-a nego u mitohondrijima. ATP se razgrađuje kidanjem veza fosfor-kiseonik i oslobađanjem energije. To se događa pod djelovanjem enzima ATPaze u procesu hidrolize ATP-a - dodavanja vode uz eliminaciju molekula fosforne kiseline. Kao rezultat toga, ATP se pretvara u ADP, a ako se odvoje dva molekula fosforne kiseline, onda u AMP. Reakcija cijepanja svakog gram-molekula kiseline je praćena oslobađanjem 40 kJ. Ovo je vrlo veliki energetski prinos, pa se fosfor-kiseoničke veze ATP-a obično nazivaju makroergijskim (visokoenergetskim).
Korištenje ATP-a u reakcijama plastične izmjene provodi se njihovom konjugacijom s hidrolizom ATP-a. Molekuli različitih supstanci se nabijaju energijom vezivanjem fosforne grupe koja se oslobađa tokom hidrolize iz molekula ATP-a, odnosno fosforilacijom.
Karakteristika fosfatnih derivata je da ne mogu napustiti ćeliju, iako njihovi "ispražnjeni" oblici slobodno prolaze kroz membranu. Zbog toga fosforilirani molekuli ostaju u ćeliji sve dok se ne iskoriste u odgovarajućim reakcijama.
Obrnuti proces pretvaranja ADP-a u ATP događa se pričvršćivanjem molekula fosforne kiseline na ADP, oslobađanjem vode i apsorbiranjem velike količine energije.
Dakle, ATP je univerzalni i neposredni izvor energije za ćelijsku aktivnost. Ovo stvara jedinstveni ćelijski fond energije i omogućava njegovu redistribuciju i transport iz jednog dijela ćelije u drugi.
Prijenos fosfatne grupe igra važnu ulogu u kemijskim reakcijama kao što je sastavljanje makromolekula iz monomera. Na primjer, aminokiseline se mogu kombinovati u peptide samo ako su prethodno fosforilirane. Mehanički procesi kontrakcije ili kretanja, prijenos otopljene tvari protiv gradijenta koncentracije i drugi procesi povezani su s trošenjem energije pohranjene u ATP-u.
Proces razmjene energije može se predstaviti na sljedeći način. Visokomolekularne organske tvari u citoplazmi se enzimski, hidrolizom, pretvaraju u jednostavnije, od kojih se sastoje: proteini - u aminokiseline, poli- i disaharidi - u monosaharide (+ glukoza), masti u glicerol i masne kiseline. Oksidativni procesi su odsutni, oslobađa se malo energije koja se ne koristi i prelazi u termalni oblik. Većina ćelija prvo koristi ugljikohidrate. Polisaharidi (škrob u biljkama i glikogen kod životinja) hidroliziraju se u glukozu. Oksidacija glukoze se odvija u tri faze: glikoliza, oksidativna dekarboksilacija (Krebsov ciklus - ciklus limunske kiseline) i oksidativna fosforilacija (respiratorni lanac). U citoplazmi se odvija glikoliza, uslijed koje se jedan molekul glukoze dijeli na dva molekula pirogrožđane kiseline uz oslobađanje dva molekula ATP-a. U nedostatku kisika, pirogrožđana kiselina se pretvara u etanol (fermentacija) ili mliječnu kiselinu (anaerobno disanje).
Ako se glikoliza provodi u životinjskim stanicama, molekula glukoze sa šest ugljika razlaže se na dva molekula mliječne kiseline. Ovaj proces je višestepeni. Izvodi ga uzastopno 13 enzima. Tokom alkoholne fermentacije, iz molekula glukoze nastaju dva molekula etanola i dva molekula CO2.
Glikoliza je faza zajednička za anaerobno i aerobno disanje, druge dvije se provode samo u aerobnim uvjetima. Proces oksidacije bez kisika, u kojem se oslobađa i koristi samo dio energije metabolita, konačni je proces za anaerobne organizme. U prisustvu kiseonika, pirogrožđana kiselina prelazi u mitohondrije, gde se kao rezultat niza uzastopnih reakcija potpuno aerobno oksidira u H2O i CO2 uz istovremenu fosforilaciju ADP-a u ATP. Istovremeno, glikoliza daje dva ATP molekula, dva - Krebsov ciklus, 34 - respiratorni lanac. Neto prinos od potpune oksidacije jednog molekula glukoze u H2O i CO2 je 38 molekula.
Tako se u aerobnim organizmima konačna razgradnja organskih tvari vrši oksidacijom atmosferskim kisikom u jednostavne anorganske tvari: CO2 i H2O. Ovaj proces se odvija na kristama mitohondrija. U tom slučaju se oslobađa maksimalna količina slobodne energije, čiji je značajan dio rezervisan u molekulima ATP-a. Lako je vidjeti da aerobna oksidacija u najvećoj mjeri daje ćeliji besplatnu energiju.
Kao rezultat katabolizma, energetski bogati ATP molekuli se akumuliraju u ćeliji, a CO2 i višak vode oslobađaju se u vanjsko okruženje.
Molekuli šećera koji nisu potrebni za disanje mogu se pohraniti u ćeliji. Višak lipida se ili cijepa, nakon čega produkti njihovog cijepanja ulaze u mitohondrije kao supstrat za disanje, ili se talože u rezervi u citoplazmi u obliku masnih kapljica. Proteini se grade od aminokiselina koje ulaze u ćeliju. Sinteza proteina se odvija u organelama zvanim ribosomi. Svaki ribosom se sastoji od dvije podčestice - velike i male: obje podčestice uključuju proteinske molekule i molekule RNK.
Ribosomi su često vezani za poseban sistem membrana, koji se sastoji od tankova i vezikula, za takozvani endoplazmatski retikulum (ER); u ćelijama koje proizvode mnogo proteina, endoplazmatski retikulum je često vrlo dobro razvijen i pun ribozoma. Neki enzimi su efikasni samo ako su vezani za membranu. Većina enzima uključenih u sintezu lipida nalazi se ovdje. Dakle, endoplazmatski retikulum je, takoreći, neka vrsta ćelijske radne površine.
Osim toga, ER dijeli citoplazmu na odvojene dijelove, odnosno odjeljke, tj. odvaja različite kemijske procese koji se istovremeno odvijaju u citoplazmi, i na taj način smanjuje vjerovatnoću da će ti procesi interferirati jedan s drugim.
Često se proizvodi formirani od strane određene ćelije koriste izvan ćelije. U takvim slučajevima, proteini sintetizirani na ribosomima prolaze kroz membrane endoplazmatskog retikuluma i spakuju se u membranske vezikule koje se formiraju oko njih, koje se zatim povezuju iz ER. Ovi mjehurići, koji se spljoštavaju i slažu jedan na drugi, poput palačinki u hrpi, formiraju karakterističnu strukturu koja se naziva Golgijev kompleks ili Golgijev aparat. Tokom boravka u Golgijevom aparatu, proteini prolaze kroz određene promjene. Kada dođe vrijeme da napuste ćeliju, membranski mjehurići se spajaju sa ćelijskom membranom i prazne, izlivajući svoj sadržaj prema van, tj. do izlučivanja dolazi egzocitozom.
Lizozomi se također formiraju u Golgijevom aparatu - membranskim vrećicama koje sadrže probavne enzime. Razumijevanje kako ćelija proizvodi, pakuje i izvozi određene proteine i kako "zna" koje proteine treba zadržati za sebe, jedna je od najfascinantnijih grana moderne citologije.
Membrane bilo koje ćelije se stalno kreću i mijenjaju. ER membrane se polako kreću kroz ćeliju. Odvojeni dijelovi ovih membrana se odvajaju i formiraju vezikule, koje privremeno postaju dio Golgijevog aparata, a zatim se u procesu egzocitoze spajaju sa ćelijskom membranom.
Kasnije se materijal membrane vraća u citoplazmu, gdje se ponovo koristi.
Razmjena supstanci koje ulaze u ćeliju ili se iz nje ispuštaju van, kao i razmjena različitih signala sa mikro i makro okruženjem, odvija se kroz vanjsku membranu ćelije. Kao što je poznato, ćelijska membrana je lipidni dvosloj u koji su ugrađeni različiti proteinski molekuli koji djeluju kao specijalizirani receptori, jonski kanali, uređaji koji aktivno prenose ili uklanjaju različite kemikalije, međućelijske kontakte itd. U zdravim eukariotskim ćelijama fosfolipidi su raspoređeni u membrana asimetrično: vanjska površina se sastoji od sfingomijelina i fosfatidilholina, unutrašnja se sastoji od fosfatidilserina i fosfatidiletanolamina. Održavanje takve asimetrije zahtijeva utrošak energije. Stoga, u slučaju oštećenja ćelije, njene infekcije, energetskog gladovanja, vanjska površina membrane je obogaćena za nju neuobičajenim fosfolipidima, što postaje signal drugim stanicama i enzimima o oštećenju stanice uz odgovarajuću reakciju na nju. Najvažniju ulogu ima rastvorljivi oblik fosfolipaze A2, koji razgrađuje arahidonsku kiselinu i stvara lizoforme od navedenih fosfolipida. Arahidonska kiselina je ograničavajuća karika za stvaranje medijatora upale kao što su eikozanoidi, a zaštitni molekuli - pentraksini (C-reaktivni protein (CRP), prekursori amiloidnih proteina) - su vezani za lizoforme u membrani, nakon čega slijedi aktivacija sistem komplementa duž klasičnog puta i destrukcije ćelija.
Struktura membrane doprinosi očuvanju karakteristika unutrašnjeg okruženja ćelije, njenih razlika od spoljašnjeg okruženja. To je osigurano selektivnom propusnošću stanične membrane, postojanjem aktivnih transportnih mehanizama u njoj. Njihovo kršenje kao rezultat izravnog oštećenja, na primjer, tetrodotoksinom, ouabainom, tetraetilamonijem, ili u slučaju nedovoljne opskrbe energijom odgovarajućih "pumpi", dovodi do kršenja sastava elektrolita ćelije, promjene u njenom metabolizmu. , kršenje specifičnih funkcija - kontrakcija, provođenje impulsa ekscitacije, itd. Povreda ćelijskih jonskih kanala (kalcijum, natrijum, kalijum i hlorid) kod ljudi takođe može biti genetski određena mutacijom gena odgovornih za strukturu ovih kanala. Takozvane kanalopatije uzrok su nasljednih bolesti nervnog, mišićnog i probavnog sistema. Prekomjeran unos vode u ćeliju može dovesti do njenog pucanja – citolize – zbog perforacije membrane tokom aktivacije komplementa ili napada citotoksičnih limfocita i prirodnih ubica.
Mnogi receptori su ugrađeni u ćelijsku membranu - strukture koje u kombinaciji sa odgovarajućim specifičnim signalnim molekulima (ligandima) prenose signal u ćeliju. To se dešava kroz različite regulatorne kaskade, koje se sastoje od enzimski aktivnih molekula, koji se sekvencijalno aktiviraju i na kraju doprinose implementaciji različitih ćelijskih programa, kao što su rast i proliferacija, diferencijacija, pokretljivost, starenje i smrt ćelije. Regulatorne kaskade su prilično brojne, ali njihov broj još nije u potpunosti utvrđen. Sistem receptora i regulatorne kaskade povezane sa njima takođe postoje unutar ćelije; stvaraju određenu regulatornu mrežu sa tačkama koncentracije, distribucije i izbora daljeg puta signala u zavisnosti od funkcionalnog stanja ćelije, faze njenog razvoja i istovremenog delovanja signala sa drugih receptora. Rezultat toga može biti inhibicija ili pojačanje signala, njegovo usmjeravanje duž drugačijeg regulatornog puta. I receptorski aparat i putevi transdukcije signala kroz regulatorne kaskade, na primjer do jedra, mogu biti poremećeni kao rezultat genetskog defekta koji se javlja kao urođeni defekt na nivou organizma ili zbog somatske mutacije u određenoj ćeliji. tip. Ovi mehanizmi mogu biti oštećeni infektivnim agensima, toksinima, a mogu se promijeniti i tokom starenja. Posljednja faza ovoga može biti kršenje funkcija ćelije, procesa njene proliferacije i diferencijacije.
Molekuli koji igraju važnu ulogu u procesima međućelijske interakcije nalaze se i na površini ćelija. To mogu uključivati proteine ćelijske adhezije, antigene kompatibilnosti tkiva, tkivno-specifične, diferencirajuće antigene itd. Promjene u sastavu ovih molekula uzrokuju kršenje međustaničnih interakcija i mogu uzrokovati aktivaciju odgovarajućih mehanizama za eliminaciju takvih ćelija, jer predstavljaju određenu opasnost po integritet organizma kao rezervoar infekcije, posebno virusne, ili kao potencijalni pokretači rasta tumora.
Kršenje opskrbe energijom ćelije
Izvor energije u ćeliji je hrana, nakon čijeg se razgradnje energija oslobađa do konačnih tvari. Mitohondrije su glavno mjesto proizvodnje energije u kojoj se tvari oksidiraju uz pomoć enzima respiratornog lanca. Oksidacija je glavni dobavljač energije, jer se kao rezultat glikolize ne oslobađa više od 5% energije iz iste količine oksidacijskih supstrata (glukoze), u odnosu na oksidaciju. Oko 60% energije koja se oslobađa tokom oksidacije akumulira se oksidativnom fosforilacijom u makroergijskim fosfatima (ATP, kreatin fosfat), ostatak se raspršuje kao toplota. U budućnosti, visokoenergetske fosfate koristi ćelija za procese kao što su pumpanje, sinteza, dioba, kretanje, sekrecija, itd. Postoje tri mehanizma čije oštećenje može uzrokovati poremećaj u opskrbi ćelije energijom. : prvi je mehanizam za sintezu enzima energetskog metabolizma, drugi je mehanizam oksidativne fosforilacije, treći - mehanizam korištenja energije.
Poremećaj transporta elektrona u respiratornom lancu mitohondrija ili odvajanje ADP oksidacije i fosforilacije sa gubitkom protonskog potencijala - pokretačke sile stvaranja ATP-a, dovodi do slabljenja oksidativne fosforilacije na način da se većina energije raspršuje u smanjuje se oblik toplote i broj makroergijskih jedinjenja. Razdvajanje oksidacije i fosforilacije pod uticajem adrenalina koriste ćelije homoiotermnih organizama za povećanje proizvodnje toplote uz održavanje konstantne telesne temperature tokom hlađenja ili njenog povećanja tokom groznice. Kod tireotoksikoze uočavaju se značajne promjene u strukturi mitohondrija i energetskom metabolizmu. Ove promjene su u početku reverzibilne, ali nakon određenog trenutka postaju ireverzibilne: mitohondrije se fragmentiraju, raspadaju ili nabubre, gube kriste, pretvaraju se u vakuole i na kraju akumuliraju tvari kao što su hijalin, feritin, kalcij, lipofuscin. Kod pacijenata sa skorbutom, mitohondrije se spajaju i formiraju hondriosfere, vjerovatno zbog oštećenja membrane peroksidnim spojevima. Značajna oštećenja mitohondrija nastaju pod uticajem jonizujućeg zračenja, prilikom transformacije normalne ćelije u malignu.
Mitohondrije su moćan depo jona kalcijuma, gde je njegova koncentracija nekoliko redova veličine veća od one u citoplazmi. Kada su mitohondriji oštećeni, kalcij ulazi u citoplazmu, uzrokujući aktivaciju proteinaza s oštećenjem intracelularnih struktura i poremećajem funkcija odgovarajuće stanice, na primjer, kalcijeve kontrakture ili čak "kalcijevu smrt" u neuronima. Kao rezultat narušavanja funkcionalne sposobnosti mitohondrija, naglo se povećava stvaranje slobodnih radikala peroksidnih spojeva, koji imaju vrlo visoku reaktivnost i stoga oštećuju važne ćelijske komponente - nukleinske kiseline, proteine i lipide. Ova pojava se uočava tokom takozvanog oksidativnog stresa i može imati negativne posledice na postojanje ćelije. Dakle, oštećenje vanjske mitohondrijalne membrane je praćeno oslobađanjem u citoplazmu tvari koje se nalaze u međumembranskom prostoru, prvenstveno citokroma C i nekih drugih biološki aktivnih tvari, koje pokreću lančane reakcije koje uzrokuju programiranu smrt stanice – apoptozu. Oštećujući mitohondrijsku DNK, reakcije slobodnih radikala iskrivljuju genetske informacije neophodne za formiranje određenih enzima respiratornog lanca koji se proizvode specifično u mitohondrijima. To dovodi do još većeg poremećaja oksidativnih procesa. U cjelini, intrinzični genetski aparat mitohondrija, u poređenju sa genetskim aparatom jezgra, manje je zaštićen od štetnih utjecaja koji mogu promijeniti genetske informacije koje su u njemu kodirane. Kao rezultat toga, mitohondrijska disfunkcija se javlja tijekom cijelog života, na primjer, u procesu starenja, tijekom maligne transformacije stanice, kao iu pozadini nasljednih mitohondrijalnih bolesti povezanih s mutacijom mitohondrijske DNK u jajetu. Trenutno je opisano više od 50 mitohondrijalnih mutacija koje uzrokuju nasljedne degenerativne bolesti nervnog i mišićnog sistema. Prenose se na dijete isključivo od majke, jer mitohondrije sperme nisu dio zigota i, shodno tome, novog organizma.
Kršenje očuvanja i prijenosa genetskih informacija
Ćelijsko jezgro sadrži većinu genetskih informacija i na taj način osigurava njegovo normalno funkcioniranje. Uz pomoć selektivne ekspresije gena koordinira rad ćelije u interfazi, pohranjuje genetske informacije, rekreira i prenosi genetski materijal u procesu diobe ćelije. Replikacija DNK i transkripcija RNK odvijaju se u jezgru. Razni patogeni faktori, kao što su ultraljubičasto i jonizujuće zračenje, oksidacija slobodnih radikala, hemikalije, virusi, mogu oštetiti DNK. Procjenjuje se da svaka ćelija toplokrvne životinje u jednom danu. gubi preko 10.000 baza. Ovome treba dodati i prekršaje prilikom kopiranja tokom podjele. Ako se ovo oštećenje nastavi, ćelija ne bi mogla preživjeti. Zaštita leži u postojanju moćnih sistema popravke, kao što je ultraljubičasta endonukleaza, sistem reparativne replikacije i rekombinacione popravke, koji zamenjuju oštećenje DNK. Genetski defekti reparativnih sistema uzrokuju razvoj bolesti zbog povećane osjetljivosti na faktore koji oštećuju DNK. Riječ je o pigmentiranoj kserodermi, kao io nekim sindromima ubrzanog starenja, praćeni povećanom sklonošću nastanku malignih tumora.
Sistem regulacije procesa replikacije DNK, transkripcije glasničke RNK (mRNA), prevođenja genetskih informacija iz nukleinskih kiselina u strukturu proteina je prilično složen i višeslojan. Pored regulatornih kaskada koje pokreću djelovanje više od 3000 faktora transkripcije koji aktiviraju određene gene, postoji i višeslojni regulatorni sistem posredovan malim RNA molekulima (interferirajuće RNA; RNAi). Ljudski genom, koji se sastoji od otprilike 3 milijarde purinskih i pirimidinskih baza, sadrži samo 2% strukturnih gena odgovornih za sintezu proteina. Ostatak osigurava sintezu regulatornih RNK, koje zajedno sa transkripcijskim faktorima aktiviraju ili blokiraju rad strukturnih gena na nivou DNK u hromozomima ili utiču na translaciju glasničke RNK (mRNA) tokom formiranja polipeptidnog molekula u citoplazmi. . Povreda genetskih informacija može se javiti kako na nivou strukturnih gena, tako i na regulatornom dijelu DNK sa odgovarajućim manifestacijama u vidu različitih nasljednih bolesti.
U posljednje vrijeme se velika pažnja poklanja promjenama u genetskom materijalu koje nastaju tijekom individualnog razvoja organizma i povezane su sa inhibicijom ili aktivacijom određenih dijelova DNK i hromozoma zbog njihove metilacije, acetilacije i fosforilacije. Ove promjene traju dugo vremena, ponekad tokom cijelog života organizma od embriogeneze do starosti, a nazivaju se epigenomskim nasljeđem.
Reprodukciju ćelija sa izmenjenom genetskom informacijom sprečavaju i sistemi (faktori) kontrole mitotičkog ciklusa. Oni stupaju u interakciju sa ciklin zavisnim protein kinazama i njihovim katalitičkim podjedinicama - ciklinima - i blokiraju prolazak kompletnog mitotičkog ciklusa u ćeliji, zaustavljajući podjelu na granici između presintetičke i sintetičke faze (blokiraju G1/S) dok se popravak DNK ne završi , a ako je to nemoguće, pokreću programirane ćelije smrti. Ovi faktori uključuju p53 gen, čija mutacija uzrokuje gubitak kontrole nad proliferacijom transformiranih ćelija; javlja se u skoro 50% slučajeva raka kod ljudi. Druga kontrolna tačka prolaska mitotičkog ciklusa nalazi se na granici G2/M. Ovdje se ispravna distribucija hromozomskog materijala između ćelija kćeri u mitozi ili mejozi kontrolira korištenjem kompleksa mehanizama koji kontroliraju ćelijsko vreteno, centar i centromere (kinetohore). Neefikasnost ovih mehanizama dovodi do narušavanja distribucije hromozoma ili njihovih delova, što se manifestuje odsustvom bilo kog hromozoma u jednoj od ćelija kćeri (aneuploidija), prisustvom dodatnog hromozoma (poliploidija), odvajanjem dio hromozoma (delecija) i njegov prijenos na drugi hromozom (translokacija). Ovakvi procesi se vrlo često uočavaju prilikom reprodukcije maligno degenerisanih i transformisanih ćelija. Ako se to dogodi tokom mejoze sa zametnim stanicama, to dovodi ili do smrti fetusa u ranoj fazi embrionalnog razvoja, ili do rođenja organizma s hromozomskom bolešću.
Nekontrolisana reprodukcija ćelija tokom rasta tumora nastaje kao rezultat mutacija u genima koji kontrolišu proliferaciju ćelija i nazivaju se onkogeni. Među više od 70 trenutno poznatih onkogena, većina njih su komponente regulacije rasta ćelija, neki su transkripcioni faktori koji regulišu aktivnost gena, kao i faktori koji inhibiraju deobu i rast ćelija. Drugi faktor koji ograničava prekomjerno širenje (širenje) proliferirajućih stanica je skraćivanje krajeva hromozoma - telomera, koji se ne mogu u potpunosti replicirati kao rezultat čisto sterične interakcije, pa se nakon svake diobe ćelije telomeri skraćuju za određeni dio baza. Tako proliferirajuće ćelije odraslog organizma nakon određenog broja dioba (obično od 20 do 100, ovisno o vrsti organizma i njegovoj starosti) iscrpljuju dužinu telomera i zaustavlja se daljnja replikacija hromozoma. Ovaj fenomen se ne javlja u spermatogenom epitelu, enterocitima i embrionalnim ćelijama zbog prisustva enzima telomeraze, koji obnavlja dužinu telomera nakon svake deobe. U većini ćelija odraslih organizama telomeraza je blokirana, ali se, nažalost, aktivira u tumorskim ćelijama.
Veza između jezgra i citoplazme, transport tvari u oba smjera odvijaju se kroz pore u nuklearnoj membrani uz učešće posebnih transportnih sistema sa potrošnjom energije. Tako se energija i plastične supstance, signalni molekuli (transkripcioni faktori) transportuju do jezgra. Obrnuti tok dovodi u citoplazmu molekule mRNA i transfer RNK (tRNA), ribozome neophodne za sintezu proteina u ćeliji. Isti način transporta supstanci je svojstven virusima, posebno, kao što je HIV. Oni prenose svoj genetski materijal u jezgro ćelije domaćina sa njegovim daljim uključivanjem u genom domaćina i prenosom novoformirane virusne RNK u citoplazmu za dalju sintezu proteina novih virusnih čestica.
Kršenje procesa sinteze
Procesi sinteze proteina odvijaju se u cisternama endoplazmatskog retikuluma, usko povezanim s porama u nuklearnoj membrani, kroz koje ribozomi, tRNA i mRNA ulaze u endoplazmatski retikulum. Ovdje se vrši sinteza polipeptidnih lanaca koji kasnije dobijaju svoj konačni oblik u agranularnom endoplazmatskom retikulumu i lamelarnom kompleksu (Golgijev kompleks), gdje se podvrgavaju posttranslacijskoj modifikaciji i povezivanju s molekulama ugljikohidrata i lipida. Novonastali proteinski molekuli ne ostaju na mjestu sinteze, već uz pomoć složenog reguliranog procesa, tzv. protein kineza, aktivno se prenose u onaj izolirani dio ćelije gdje će obavljati svoju predviđenu funkciju. U ovom slučaju, vrlo važan korak je strukturiranje prenesenog molekula u odgovarajuću prostornu konfiguraciju sposobnu da obavlja svoju inherentnu funkciju. Takvo strukturiranje se dešava uz pomoć posebnih enzima ili na matrici specijalizovanih proteinskih molekula - šaperona, koji pomažu da molekul proteina, novoformiran ili izmenjen usled spoljašnjeg uticaja, dobije ispravnu trodimenzionalnu strukturu. U slučaju štetnog djelovanja na ćeliju, kada postoji mogućnost kršenja strukture proteinskih molekula (na primjer, s povećanjem tjelesne temperature, infektivnim procesom, intoksikacijom), koncentracija pratioca u ćeliji naglo raste. Stoga se takvi molekuli još nazivaju proteini stresa, ili proteini toplotnog šoka. Povreda strukturiranja proteinske molekule dovodi do stvaranja hemijski inertnih konglomerata koji se talože u ćeliji ili izvan nje u slučaju amiloidoze, Alchajmerove bolesti itd. će biti defektni. Ova situacija se javlja kod tzv. prionskih bolesti (scrappie kod ovaca, bjesnoća kod krava, kuru, Creutzfeldt-Jakobova bolest kod ljudi), kada defekt u jednom od membranskih proteina nervne ćelije uzrokuje naknadno nakupljanje inertnih masa u unutrašnjosti. ćelije i poremećaj njene vitalne aktivnosti.
Do kršenja procesa sinteze u ćeliji može doći u različitim fazama: transkripcija RNK u jezgru, translacija polipeptida u ribosomima, posttranslacijska modifikacija, hipermetilacija i glikozilacija bež molekula, transport i distribucija proteina u ćeliji i njihovo uklanjanje spolja. U ovom slučaju može se uočiti povećanje ili smanjenje broja ribozoma, raspad poliribosoma, širenje cisterni granularnog endoplazmatskog retikuluma, gubitak ribozoma njime, stvaranje vezikula i vakuola. Dakle, u slučaju trovanja blijedom žabokrečinom dolazi do oštećenja enzima RNA polimeraze, što remeti transkripciju. Toksin difterije, inaktivirajući faktor elongacije, ometa procese translacije, uzrokujući oštećenje miokarda. Razlog za kršenje sinteze nekih specifičnih proteinskih molekula mogu biti infektivni agensi. Na primjer, herpesvirusi inhibiraju sintezu i ekspresiju molekula MHC antigena, što im omogućava da djelimično izbjegnu imunološku kontrolu, a bacili kuge inhibiraju sintezu medijatora akutne upale. Pojava neobičnih proteina može zaustaviti njihov daljnji razgradnju i dovesti do nakupljanja inertnog ili čak toksičnog materijala. Tome u određenoj mjeri može doprinijeti i poremećaj procesa raspadanja.
Kršenje procesa propadanja
Istovremeno sa sintezom proteina u ćeliji, kontinuirano dolazi do njenog propadanja. U normalnim uslovima, ovo ima važan regulatorni i formativni značaj, na primer, tokom aktivacije neaktivnih oblika enzima, proteinskih hormona i proteina mitotičkog ciklusa. Normalan rast i razvoj ćelije zahtevaju fino kontrolisanu ravnotežu između sinteze i razgradnje proteina i organela. Međutim, u procesu sinteze proteina, zbog grešaka u radu aparata za sintezu, abnormalnog strukturiranja proteinske molekule, njenog oštećenja kemijskim i bakterijskim agensima, konstantno se stvara prilično veliki broj neispravnih molekula. Prema nekim procjenama, njihov udio je oko trećine svih sintetiziranih proteina.
Ćelije sisara imaju nekoliko glavnih putevi razgradnje proteina: kroz lizozomalne proteaze (pentidne hidrolaze), proteinaze zavisne od kalcijuma (endopeptidaze) i proteazomski sistem. Osim toga, postoje i specijalizirane proteinaze, kao što su kaspaze. Glavna organela u kojoj dolazi do razgradnje tvari u eukariotskim stanicama je lizozom, koji sadrži brojne hidrolitičke enzime. Usljed procesa endocitoze i raznih tipova autofagije u lizosomima i fagolizosomima uništavaju se kako defektni proteinski molekuli tako i cijele organele: oštećeni mitohondriji, dijelovi plazma membrane, neki ekstracelularni proteini, sadržaj sekretornih granula.
Važan mehanizam razgradnje proteina je proteasom, složena multikatalitička proteinazna struktura lokalizovana u citosolu, jezgru, endoplazmatskom retikulumu i na ćelijskoj membrani. Ovaj enzimski sistem je odgovoran za razgradnju oštećenih proteina, kao i zdravih proteina koji se moraju ukloniti za normalnu funkciju ćelija. U ovom slučaju, proteini koji se uništavaju se preliminarno kombinuju sa specifičnim polipeptidom ubikvitina. Međutim, proteini koji nisu ubikvitirani također mogu biti djelomično uništeni u proteazomima. Razgradnja proteinske molekule u proteazomima do kratkih polipeptida (obrada) sa njihovom naknadnom prezentacijom zajedno sa molekulima MHC tipa I važna je karika u sprovođenju imunološke kontrole antigenske homeostaze organizma. Kada je funkcija proteasoma oslabljena, dolazi do nakupljanja oštećenih i nepotrebnih proteina, što prati starenje stanica. Kršenje razgradnje proteina ovisnih o ciklinu dovodi do kršenja diobe stanica, degradacije sekretornih proteina - do razvoja cistofibroze. Suprotno tome, povećanje funkcije proteasoma prati iscrpljivanje organizma (AIDS, rak).
Uz genetski uvjetovane poremećaje degradacije proteina, organizam nije održiv i umire u ranim fazama embriogeneze. Ako je poremećena razgradnja masti ili ugljikohidrata, dolazi do bolesti nakupljanja (tezaurizmoze). Istovremeno se unutar ćelije nakuplja prekomjerna količina određenih tvari ili proizvoda njihove nepotpune razgradnje – lipida, polisaharida, što značajno narušava funkciju stanice. Najčešće se opaža u epiteliocitima jetre (hepatocitima), neuronima, fibroblastima i makrofagocitima.
Stečeni poremećaji u procesima raspadanja tvari mogu nastati kao posljedica patoloških procesa (na primjer, proteinska, masna, ugljikohidratna i pigmentna distrofija) i biti praćeni stvaranjem neobičnih tvari. Poremećaji u sistemu lizosomske proteolize dovode do smanjenja adaptacije tokom gladovanja ili povećanog opterećenja, do pojave nekih endokrinih disfunkcija - smanjenja nivoa insulina, tireoglobulina, citokina i njihovih receptora. Poremećaji razgradnje proteina usporavaju brzinu zacjeljivanja rana, uzrokuju razvoj ateroskleroze i utiču na imunološki odgovor. U slučaju hipoksije, promjene intracelularnog pH, ozljede zračenja, koje karakterizira povećana peroksidacija membranskih lipida, kao i pod utjecajem lizosomotropnih supstanci - endotoksina bakterija, metabolita toksičnih gljivica (sporofusarin), kristala silicijum oksida - stabilnost membrane lizosoma promjene, aktivirani lizosomski enzimi se oslobađaju u citoplazmu, što uzrokuje uništavanje staničnih struktura i njihovu smrt.
Poglavlje 1
OSNOVE FIZIOLOGIJE ĆELIJE
I. Dudel
plazma membrana . Životinjske ćelije su ograničene plazma membranom (slika 1.1). Detaljnije ćemo se zadržati na njegovoj strukturi, koja je vrlo slična strukturi mnogih intracelularnih membrana. Glavna matrica membrane se sastoji od lipida uglavnom fosfatidilholin. Ovi lipidi se sastoje od hidrofilne glavne grupe za koju su vezani dugi hidrofobni ugljikovodični lanci. U vodi takvi lipidi spontano formiraju dvoslojni film debljine 4-5 nm, u kojem su hidrofilne grupe okrenute prema vodenom mediju, a hidrofobni ugljikovodični lanci su raspoređeni u dva reda, formirajući bezvodnu lipidnu fazu. Ćelijske membrane su lipidni dvoslojevi ovog tipa i sadrže glikolipide, holesterol i fosfolipide (slika 1.2). Hidrofilni dio glikolipida čine oligosaharidi. Glikolipidi se uvijek nalaze na vanjskoj površini plazma membrane, a oligosaharidni dio molekule orijentiran je poput dlake uronjene u okolinu. Rasuti među fosfolipidima u skoro jednakim količinama molekula holesterola stabilizuju membranu. Raspodjela različitih lipida u unutrašnjem i vanjskom sloju membrane nije ista, a čak i unutar istog sloja postoje područja u kojima su koncentrisane određene vrste lipida. Tako neravnomjerna raspodjela
Rice. 1.1. Šematski crtež ćelije koji prikazuje najvažnije organele
vjerovatno ima neki, još nejasan, funkcionalni značaj.
Glavni funkcionalni elementi uronjeni u relativno inertni lipidni matriks membrane su vjeverice(Sl. 1.2). Protein po težini se kreće od 25 do 75% u različitim membranama, ali budući da su proteinski molekuli mnogo veći od molekula lipida, 50% mase je ekvivalentno odnosu od 1 proteinske molekule prema 50 molekula lipida. Neki proteini prodiru kroz membranu od njene vanjske do unutrašnje površine, dok su drugi fiksirani u jednom sloju. Proteinski molekuli su obično orijentisani tako da su njihove hidrofobne grupe uronjene u lipidnu membranu, a polarne hidrofilne grupe na površini membrane uronjene u vodenu fazu. Mnogi proteini vanjske površine membrane su glikoproteini; njihove hidrofilne saharidne grupe su okrenute vanćelijskom okruženju.
Membranski sistemi intracelularnih organela .
Otprilike polovinu volumena ćelije zauzimaju organele izolovane iz citosola membranama. Ukupna površina membrana intracelularnih organela je najmanje 10 puta veća od površine plazma membrane. Najrasprostranjeniji je membranski sistem endoplazmatski retikulum, predstavlja mrežu

Rice. 1.2.Šematski prikaz plazma membrane. Proteini su ugrađeni u fosfolipidni dvosloj, pri čemu neki pokrivaju dvosloj dok su drugi samo usidreni za vanjski ili unutrašnji sloj
visoko izvijene tubule ili vrećaste izdužene strukture; velike površine endoplazmatskog retikuluma su prošarane ribosomima; takav retikulum se naziva granular, ili hrapav (slika 1.1). golgijev aparat takođe se sastoji od membranom vezanih lamela, od kojih se odvajaju vezikule, ili vezikule (slika 1.1). Lizozomi i peroksizomi su male specijalizovane vezikule. U svim ovim raznolikim organelama, membrana i prostor koji pokriva sadrže specifične skupove enzima; unutar organela se nakupljaju posebni metabolički produkti koji služe za obavljanje različitih funkcija organela.
CoreI mitohondrije razlikuju se po tome što je svaka od ovih organela okružena s dvije membrane. Jezgro je odgovorno za kinetičku kontrolu metabolizma; presavijena unutrašnja mitohondrijalna membrana je mjesto oksidativnog metabolizma; ovdje se, zbog oksidacije piruvata ili masnih kiselina, sintetiše visokoenergetski spoj adenozin trifosfat (ATP, ili ATP).
citoskelet . Citoplazma koja okružuje organele nikako se ne može smatrati amorfnom; prožeta je mrežom citoskeleta. Citoskelet se sastoji od mikrotubula, aktinskih filamenata i intermedijarnih filamenata (slika 1.1). mikrotubule imaju vanjski prečnik od oko 25 nm; nastaju, poput normalnog polimera, kao rezultat sklapanja proteinskih molekula tubulina. aktinski filamenti- kontraktilna vlakna koja se nalaze u sloju blizu membrane i kroz celu ćeliju - uglavnom učestvuju u procesima povezanim sa kretanjem. Intermedijarni filamenti sastoje se od blokova različitog hemijskog sastava u različitim tipovima ćelija; oni formiraju razne veze između druga dva elementa citoskeleta pomenuta gore. Organele i plazma membrana su također povezane sa citoskeletom, koji ne samo da održava oblik ćelije i položaj organela u njoj, već određuje i promjenu oblika ćelije i njenu pokretljivost.
Cytosol . Otprilike polovinu volumena ćelije zauzima citosol. Budući da ima otprilike 20% (težinski) proteina, više je gel nego vodeni rastvor. Male molekule, uključujući organske i neorganske joni, rastvoren u vodenoj fazi. Između ćelije i okoline (ekstracelularnog prostora) dolazi do razmene jona; o ovim procesima razmjene će biti riječi u sljedećem odjeljku. Koncentracija jona u ekstracelularnom prostoru održava se sa značajnom tačnošću na konstantnom nivou; intracelularna koncentracija svakog od jona takođe ima specifičan nivo koji se razlikuje od onog izvan ćelije (tabela 1.1). Najčešći kation u vanćelijskom okruženju je Na+ u ćeliji njegova koncentracija je više od 10 puta niža. Naprotiv, unutar ćelije koncentracija K+ je najveća, van ćelije je niža za više od reda veličine. Najveći gradijent između ekstracelularnih i intracelularnih koncentracija postoji za Ca 2+, čija je koncentracija slobodnih jona unutar ćelije najmanje 10 000 puta manja nego izvan nje. Nisu svi ioni otopljeni u citosolu; neki od njih se adsorbuju na proteinima ili deponuju u organelama. Na primjer, u slučaju Ca 2+ vezani ioni su mnogo brojniji od slobodnih. Većina proteina citosola su enzimi, uz učešće kojih se odvijaju mnogi procesi srednjeg metabolizma: glikoliza i glukoneogeneza, sinteza ili uništavanje aminokiselina, sinteza proteina na ribosomima (slika 1.1). Citosol sadrži i lipidne kapljice i granule glikogena, koje služe kao rezerve važnih molekula.
Tabela 1.1.Intra- i ekstracelularne koncentracije jona u mišićnim ćelijama homoiotermnih životinja. ALI – – “ćelijski anjoni visoke molekularne težine”
|
Intracelularna koncentracija |
Ekstracelularna koncentracija |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
sa l - |
4 mmol/l |
C1 - |
120 mmol/l |
|
HCO 3 - |
8 mmol/l |
HCO 3 - |
27 mmol/l |
|
A- |
155 mmol/l |
Ostalo katjoni |
5 mmol/l |
|
Potencijal mirovanja -90 mV |
|||
1.2. Razmjena tvari između ćelije i okoline
Ukratko smo opisali strukturu ćelije kako bismo koristili ovaj opis za pregled osnova fiziologije ćelije. Ćelija se ni u kom slučaju ne može smatrati statičkom formacijom, jer postoji stalna razmjena tvari između različitih intracelularnih odjeljaka, kao i između odjeljaka i okoline. Strukture ćelije su u dinamičkoj ravnoteži, a interakcija ćelija među sobom i sa spoljašnjom sredinom je neophodan uslov za održavanje života organizma koji funkcioniše. U ovom poglavlju ćemo razmotriti osnovne mehanizme takve razmjene. U kasnijim poglavljima, ovi mehanizmi će se razmatrati u odnosu na nervnu ćeliju i njene funkcije;
međutim, isti mehanizmi su u osnovi funkcionisanja svih drugih organa.
Difuzija.Najjednostavniji proces kretanja tvari je difuzija. U otopinama (ili plinovima), atomi i molekuli se kreću slobodno, a razlika u koncentracijama je uravnotežena difuzijom. Razmotrimo dvije zapremine ispunjene tekućinom ili plinom (slika 1.3), u kojima tvari imaju koncentracije c1 i c2 i odvojeni slojem površine A i debljine d. Protok materije m u vremenu t opisano Fikov prvi zakon difuzije:
dm/ dt= DA/ d ( C 1 –S 2)=DA/ dD C(1)
gdje je D koeficijent difuzije, koji je konstantan za datu supstancu, rastvarač i temperaturu. U opštijem obliku, za razliku koncentracije dc na udaljenosti dx
dm/dt= -D A dc/dx,(2)
protok kroz dio A je proporcionalan gradijentu koncentracije dc/dx . Znak minus se pojavljuje u jednadžbi jer je promjena koncentracije u smjeru x negativna.
Difuzija je najvažniji proces kojim se većina molekula u vodenim otopinama kreće na kratke udaljenosti. Ovo se odnosi i na njihovo kretanje u ćeliji ukoliko membrane ne ometaju difuziju. Mnoge tvari mogu slobodno difundirati kroz lipidne membrane, posebno vodu i otopljene plinove kao što su O 2 i CO 2 . rastvorljiv u mastima

Rice. 1.3.Kvantitativna shema difuzije. Dva prostora su odvojena slojem debljinedi područje ALI. C; - visoka koncentracija čestica u lijevom dijelu zapremine, C:, - niska koncentracija čestica u desnom dijelovi, roze površina je gradijent koncentracije u difuzijskom sloju. Difuzijski tok dm/dt – vidi jednadžba (1)
supstance takođe dobro difunduju kroz membrane; ovo se odnosi i na relativno male polarne molekule, kao što su etanol i urea, dok šećeri teško prolaze kroz lipidni sloj. U isto vrijeme, lipidni slojevi su praktički nepropusni za nabijene molekule, uključujući čak i neorganske ione. Za neelektrolite, jednačina difuzije (1) se obično transformiše kombinovanjem karakteristika membrane i difuzne supstance u jednu parametar-propusnost (P):
dm/dt=P AD c.(3)
Na sl. 1.4 u poređenju permeabilnost (P) lipidne membrane za različite molekule.
Difuzija kroz pore membrane . Plazma membrana (i druge ćelijske membrane) su propusne ne samo za supstance koje difundiraju kroz lipidni sloj, već i za mnoge jone, šećere, aminokiseline i nukleotide. Ove supstance prolaze kroz membranu kroz pore koje formiraju transportni proteini, ugrađen u membranu. Unutar takvih proteina postoji kanal ispunjen vodom prečnika manjeg od 1 nm kroz koji mali molekuli mogu difundirati. Kreću se duž gradijenta koncentracije, a ako nose naboj, onda je njihovo kretanje kroz kanale također regulirano membranskim potencijalom. Membranski kanali su relativno selektivni

Rice. 1.4.Propustljivost umjetnih lipidnih dvoslojeva za različite tvari
u odnosu na vrstu molekula koji kroz njih mogu proći, postoje, na primjer, kalijum, natrijum i kalcijum kanali, od kojih je svaki nepropustan za skoro sve jone osim za određeni. Takve selektivnost zbog naboja ili strukture veznih mjesta u zidovima kanala, što olakšava transport određenog molekula i sprječava prodiranje drugih tvari kroz kanal (Sl. 1.5, A) .
Iza ponašanja membranski jonski kanali Lako je zapaziti, jer se struja koja nastaje tokom kretanja jona može izmjeriti, pa čak i za jedan kanal. Pokazano je da kanali spontano i sa visokom frekvencijom mijenjaju svoje stanje iz otvorenog u zatvoreno. Kalijumski kanal karakterišu strujni impulsi sa amplitudom od oko 2 pA (2 10 -12 A) i trajanjem od nekoliko milisekundi (vidi sliku 2.12, str. 37) [3]. Tokom ovog perioda kroz njega prođu desetine hiljada jona. Prijelaz proteina iz jedne konformacije u drugu proučava se rendgenskom difrakcijom, Mössbauerovom spektroskopijom i nuklearnom magnetnom rezonancom (NMR). Proteini su stoga vrlo dinamične, pokretne strukture, a kanal kroz protein nije samo kruta cijev ispunjena vodom (slika 1.5A) već labirint molekularnih grupa i naboja koji se brzo kreću. Ovaj dinamički odgovor kanala se odražava u energetski profil kanala, prikazano na sl. 1.5, B. Ovdje apscisa prikazuje dužinu kanala od vanjskog rastvora sa koncentracijom jona C 0 i potencijalom 0 do unutrašnjeg rastvora sa koncentracijom C 1 i potencijalom E. y-osa

Rice. 1.5.A. Šema proteina koji formira kalijum kanal ugrađen u lipidni dvosloj plazma membrane. Četiri negativna naboja su fiksirana na "zidu" kanala. B. Šematski energetski profil kanala prikazanog na sl. A. Na y-osi su ucrtane vrijednosti kinetičke energije potrebne za prolazak kanala; duž ose apscise, udaljenosti između unutrašnje i vanjske površine membrane. Energetski minimumi odgovaraju mjestima vezivanja pozitivno nabijenih jona sa fiksnim negativnim nabojem u zidu kanala. Energetski maksimumi odgovaraju difuzijskim preprekama u kanalu. Pretpostavlja se da konformacija proteina kanala spontano oscilira; opcije za energetske profile su prikazane punim i isprekidanim linijama; ove oscilacije uvelike olakšavaju vezivanje jona pri savladavanju energetske barijere (ali sa promjenama)
prikazani su energetski nivoi jona na mestima vezivanja kanala; vrh na grafikonu predstavlja barijeru permeabilnosti koju energija jona mora savladati da bi prodrla u kanal, a “dip” grafikona predstavlja relativno stabilno stanje (vezivanje). Uprkos opstrukciji energetskog vrha, jon može prodrijeti u kanal ako se energetski profil spontano vrti; jon se stoga može iznenada pojaviti "s druge strane" energetskog vrha i može nastaviti da se kreće u ćeliju. U zavisnosti od naboja, veličine i stepena hidratacije jona i njegove sposobnosti da se veže za strukture zidova kanala, energetski profil kanala varira za različite jone, što može objasniti selektivnost pojedinih tipova kanala.
Difuzijska ravnoteža jona . Difuzija različitih jona kroz membranske kanale treba da dovede do eliminacije razlika u koncentracijama između ekstra- i intracelularnog okruženja. Međutim, kao što se može vidjeti iz tab. 1.1, takve razlike i dalje postoje, pa ih mora postojati ravnoteža između difuzije i drugih transportnih procesa kroz membranu. Sljedeća dva odjeljka se bave načinima na koje se uspostavlja takva ravnoteža. U slučaju jona, na difuzijsku ravnotežu utiče njihov naboj. Difuziju nenabijenih molekula osigurava razlika koncentracije dc , a kada se koncentracije izjednače, stvarni transport prestaje. Na nabijene čestice dodatno djeluje električno polje. Na primjer, kada kalijev ion izađe iz ćelije duž gradijenta koncentracije, on nosi jedan pozitivan naboj. Dakle, unutarćelijska okolina postaje negativnije nabijena, što rezultira razlikom potencijala kroz membranu. Intracelularni negativni naboj sprječava da novi ioni kalija napuste ćeliju, a oni ioni koji ipak napuste ćeliju dodatno će povećati naboj na membrani. Protok kalijevih jona prestaje kada djelovanje električnog polja kompenzira difuzijski tlak zbog razlike u koncentracijama. Joni nastavljaju da prolaze kroz membranu, ali u jednakim količinama u oba smjera. Dakle, za datu razliku u koncentraciji jona na membrani, postoji ravnotežni potencijal E ion pri čemu se zaustavlja protok jona kroz membranu. Potencijal ravnoteže može se lako odrediti pomoću Nernstove jednadžbe:
Eion= RT/ zF* lnVan/ C in(4)
gdje je R je plinska konstanta, T je apsolutna temperatura, z je valencija jona (negativna za anione) Van je ekstracelularna koncentracija jona, C in je intracelularna koncentracija jona, F Faradejev broj. Ako u jednačinu zamijenimo konstante, tada na temperaturi tijela (T = 310 K) ravnotežni potencijal za kalijeve ione E K je jednako:
Ek= –61 mB log /(5)
Ako [ K + izlaz ]/[ K + ulaz ] = 39, kako slijedi iz tabele. 1.1, dakle
Ek= -61 m B log 39= -97 mV.
Zaista, ustanovljeno je da sve ćelije imaju membranski potencijal; u mišićnim ćelijama sisara njegov nivo je oko -90 mV. U zavisnosti od uslova i relativne koncentracije jona, ćelije mogu imati membranski potencijal u rasponu od -40 do -120mV. Za ćeliju u gornjem primjeru (tabela 1.1) potencijal odmora, jednak približno -90 mV, ukazuje da su tokovi jona kalijuma kroz membranske kanale približno u ravnoteži. To nije iznenađujuće, budući da je najvjerovatnije otvoreno stanje kalijevih kanala u membrani u mirovanju; membrana je najpropusnija za jone kalija. Membranski potencijal je, međutim, određen i tokovima drugih jona.
Lakoća s kojom nenabijene čestice mogu difundirati kroz membranu kvantificirana je u jednačini (3). Propustljivost za nabijene čestice opisano malo složenijom jednadžbom:
P= m RT/ dF(6)
gdje mje pokretljivost jona u membrani, d – debljina membrane, a R, T i F poznate termodinamičke konstante. Vrijednosti permeabilnosti za različite ione određene na ovaj način mogu se koristiti za izračunavanje membranskog potencijala Em kada ioni kalijuma, natrijuma i hlorida istovremeno prolaze kroz membranu (sa propusnošću P K, P Na i P Cl odnosno). Pretpostavlja se da potencijal pada jednoliko u membrani, tako da je jačina polja konstantna. U ovom slučaju se primjenjuje Goldmanova jednačina, ili jednačina konstantnog polja :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Za većinu ćelijskih membrana P K oko 30 puta veći od R N / A (vidi također odjeljak 1.3). Relativna vrijednost PCl jako varira; za mnoge membrane PCl mali u poređenju sa R K , međutim za druge (npr. u skeletnim mišićima) PCl , mnogo veći od R K.
Aktivni transport, natrijum pumpa . Prethodni dio opisuje pasivnu difuziju jona i rezultirajući membranski potencijal pri datim intra- i ekstracelularnim koncentracijama jona. Međutim, kao rezultat ovog procesa, koncentracija jona unutar ćelije se ne stabilizuje automatski, jer membrana
potencijal je malo elektronegativniji od E K , i mnogo više u poređenju sa E N / A (oko +60 mV). Zbog difuzije, unutarćelijske koncentracije jona, barem kalija i natrijuma, trebale bi se izjednačiti sa ekstracelularnim. Stabilnost gradijenta jona postiže se aktivnim transportom: membranski proteini transportuju ione kroz membranu protiv električnih i (ili) koncentracijskih gradijenta, trošeći metaboličku energiju za to. Najvažniji proces aktivnog transporta je rad Na/K - pumpa koja postoji u skoro svim ćelijama;
pumpa pumpa jone natrijuma iz ćelije dok istovremeno pumpa jone kalijuma u ćeliju. Ovo osigurava nisku unutarćelijsku koncentraciju jona natrijuma i visoku količinu kalija (tabela 1.1). Gradijent koncentracije natrijevih jona na membrani ima specifične funkcije povezane s prijenosom informacija u obliku električnih impulsa (vidi odjeljak 2.2), kao i sa održavanjem drugih aktivnih transportnih mehanizama i regulacije volumena ćelije (vidi dolje). Stoga ne čudi da se više od 1/3 energije koju troši ćelija troši na Na/K pumpu, a u nekim od najaktivnijih ćelija i do 70% energije se troši na njen rad.
Na/K transportni protein je ATPaza. Na unutrašnjoj površini membrane razlaže ATP na ADP i fosfat (slika 1.6). Energija jednog molekula ATP-a koristi se za transport tri jona natrijuma iz ćelije i istovremeno dva jona kalija u ćeliju, odnosno ukupno se u jednom ciklusu iz ćelije ukloni jedno pozitivno naelektrisanje. Tako je Na/K pumpa elektrogeni(stvara električnu struju kroz membranu), što dovodi do povećanja elektronegativnosti membranskog potencijala za približno 10 mV. Transportni protein obavlja ovu operaciju velikom brzinom: od 150 do 600 jona natrijuma u sekundi. Aminokiselinska sekvenca transportnog proteina je poznata, ali mehanizam ovog kompleksnog razmjenskog transporta još nije jasan. Ovaj proces je opisan korišćenjem energetskih profila prenosa jona natrijuma ili kalija proteinima (slika 1.5.5). Po prirodi promene ovih profila, povezanih sa stalnim promenama u konformaciji transportnog proteina (proces koji zahteva energiju), može se suditi o stehiometriji razmene: dva jona kalijuma se zamenjuju za tri jona natrijuma.
Na/K-pumpa, kao izolovana Na+ /K+-zavisna membranska ATPaza, specifično inhibirana srčanim glikozidom ouabainom (strofantin). Pošto je rad Na/K pumpe višestepena hemijska reakcija, ona, kao i sve hemijske reakcije, u velikoj meri zavisi od temperature, što

Rice. 1.6.Šema Na/K-pumpe-ATPaze (uronjene u lipidni dvosloj plazma membrane), koja u jednom ciklusu iz ćelije protiv gradijenta potencijala i koncentracije unosi dva K iona u ćeliju + . Tokom ovog procesa, jedan ATP molekul se dijeli na ADP i fosfat. Na dijagramu, ATPaza je prikazana kao dimer koji se sastoji od velike (funkcionalne) i male podjedinice; u membrani postoji kao tetramer formiran od dvije velike i dvije male podjedinice
prikazano na sl. 1.7. Ovdje je prikazan protok jona natrijuma iz mišićnih ćelija u odnosu na vrijeme; ovo je praktično ekvivalentno protoku jona natrijuma posredovanom radom Na/K pumpe, jer je pasivni protok jona natrijuma u odnosu na koncentraciju i potencijalne gradijente izuzetno mali. Ako se preparat ohladi za oko 18°C, tada će se protok jona natrijuma iz ćelije brzo smanjiti za 15 puta, a odmah nakon zagrijavanja će se vratiti na prvobitni nivo. Ovakvo smanjenje protoka jona natrijuma iz ćelije je nekoliko puta veće od onog koji bi odgovarao temperaturnoj zavisnosti procesa difuzije ili jednostavne hemijske reakcije. Sličan efekat se primećuje kada je metabolička energija iscrpljena kao rezultat trovanja dinitrofenolom (DNP) (slika 1.7.5). Stoga se protok natrijevih jona iz ćelije osigurava reakcijom ovisnom o energiji - aktivnom pumpom. Još jedna karakteristika pumpe, uz značajnu temperaturnu i energetsku zavisnost, je prisustvo nivoa zasićenja (kao i kod svih drugih hemijskih reakcija); to znači da se brzina pumpe ne može neograničeno povećavati kako koncentracija transportiranih jona raste (slika 1.8). Nasuprot tome, protok pasivno difuzne supstance raste proporcionalno razlici u koncentracijama u skladu sa zakonom difuzije (jednadžbe 1 i 2).

Rice. 1.7. A, B. Na aktivan transport + . Y-osa: protok radioaktivnog 24 Na+ iz ćelije (imp./min). apscisa osa: vremena od početka eksperimenta. ALI.Ćelija se hladi sa 18,3°C na 0,5°C; protok Na+ izlazak iz ćelije tokom ovog perioda je inhibiran. B. Suzbijanje protoka Na+ iz ćelije dinitrofenolom (DNF) u koncentraciji od 0,2 mmol/l (sa dopunama)
Pored Na/K pumpe, plazma membrana sadrži još najmanje jednu pumpu - kalcijum; ova pumpa ispumpava jone kalcijuma (Ca 2+) iz ćelije i učestvuje u održavanju njihove intracelularne koncentracije na izuzetno niskom nivou (tabela 1.1). Kalcijum pumpa je prisutna u veoma velikoj gustini u sarkoplazmatskom retikulumu mišićnih ćelija, koje akumuliraju jone kalcijuma kao rezultat razgradnje molekula ATP (vidi Poglavlje 4).
Učinci Na/K pumpe na membranski potencijal i volumen ćelije . Na sl. 1.9 prikazuje različite komponente membranske struje i prikazuje unutarćelijske koncentracije jona koji

Rice. 1.8.Odnos između brzine transporta molekula i njihove koncentracije (na mestu ulaska u kanal ili na mestu vezivanja pumpe) tokom difuzije kroz kanal ili tokom transporta pumpom. Potonji se zasićuje pri visokim koncentracijama (maksimalna brzina, Vmax ) vrijednost na apscisi koja odgovara polovini maksimalne brzine pumpe ( Vmax /2), je ravnotežna koncentracija TO m

Rice. 1.9.Dijagram koji prikazuje koncentracije Na+ , K+ i Cl- unutar i izvan ćelije i načini na koji ti joni prodiru u ćelijsku membranu (kroz specifične jonske kanale ili uz pomoć Na/K pumpe. Sa datim gradijentima koncentracije, ravnotežni potencijali E Na, E K i E C l - jednaki su navedenim, membranski potencijal Em = – 90 mV
osigurati njihovu egzistenciju. Izlazna struja kalijevih jona se opaža kroz kalijumove kanale, budući da je membranski potencijal nešto elektropozitivniji od ravnotežnog potencijala za jone kalijuma. Ukupna provodljivost natrijumskih kanala je mnogo niža od provodljivosti kalijumovih kanala; natrijumski kanali su otvoreni mnogo rjeđe od kalijumovih kanala u stanju mirovanja; međutim, otprilike isti broj jona natrijuma ulazi u ćeliju kao i joni kalija iz nje, jer su potrebne velike koncentracije i potencijalni gradijenti za difuziju jona natrijuma u ćeliju. Na/K pumpa pruža idealnu kompenzaciju za pasivne difuzijske struje, jer transportuje ione natrijuma iz ćelije i jone kalijuma u nju. Dakle, pumpa je elektrogena zbog razlike u broju naelektrisanja prenesenih u ćeliju i iz nje, koja pri normalnoj brzini svog rada stvara membranski potencijal od oko 10 mV je elektronegativniji nego da je generiran samo pasivnim jonskim strujama (vidi jednačinu 7). Kao rezultat toga, membranski potencijal se približava potencijalu ravnoteže kalija, što smanjuje curenje jona kalija. Na aktivnost/K-pumpa se reguliše intracelularnom koncentracijom jona natrijuma. Brzina pumpe se usporava kako se smanjuje koncentracija natrijevih jona koji se uklanjaju iz ćelije (slika 1.8), tako da rad pumpe i protok natrijevih jona u ćeliju uravnotežuju jedan drugog, održavajući unutarćelijsku koncentraciju jona natrijuma na nivou od oko 10 mmol/l.
Da bi se održala ravnoteža između pumpanja i pasivnih membranskih struja, potrebno je mnogo više molekula Na/K pumpe nego što postoji kanalni protein za jone kalija i natrija. Kada je kanal otvoren, desetine hiljada jona prođu kroz njega za nekoliko milisekundi (vidi gore), a pošto se kanal obično otvara nekoliko puta u sekundi, za to vrijeme kroz njega prođe ukupno više od 10 5 jona. Jedan protein pumpe pokreće nekoliko stotina jona natrijuma u sekundi, tako da plazma membrana mora sadržavati oko 1000 puta više molekula pumpe od molekula kanala. Mjerenja strujanja kanala u mirovanju pokazala su u prosjeku jedan kalijum i jedan natrijum otvoren kanal po 1 µm 2 membrani; iz ovoga proizilazi da bi u istom prostoru trebalo biti prisutno oko 1000 Na/K molekula pumpe, tj. udaljenost između njih je u prosjeku 34 nm; prečnik pumpajućeg proteina kao proteina kanala je 8-10 nm. Dakle, membrana je dovoljno gusto zasićena pumpajućim molekulima
Činjenica da se protok jona natrijuma u ćeliju, a jona kalijuma iz ćelije kompenzuje radom pumpe, ima još jednu posledicu koja se sastoji u održavanju stabilnog osmotskog pritiska i konstantan volumen. Unutar ćelije postoji visoka koncentracija velikih anjona, uglavnom proteina (A - u tabeli 1.1), koji nisu u stanju da prodru kroz membranu (ili prodiru u nju vrlo sporo) i stoga su fiksna komponenta unutar ćelije. Da bi se uravnotežio naboj ovih anjona, potreban je jednak broj kationa. Zbog djelovanja Na/K pumpe, ovi kationi su uglavnom joni kalija. Značajno povećanje intracelularne koncentracije jona moglo bi se desiti samo sa povećanjem koncentracije anjona zbog protoka C1 - duž gradijenta koncentracije u ćeliju (tabela 1.1), ali membranski potencijal to suprotstavlja. Dolazna struja Cl- posmatra se samo dok se ne postigne ravnotežni potencijal za hloridne jone; ovo se primećuje kada je gradijent hloridnih jona skoro suprotan gradijentu kalijumovih jona, pošto su hloridni joni negativno naelektrisani (jednačina 4). Tako se uspostavlja niska intracelularna koncentracija hloridnih jona, što odgovara niskoj ekstracelularnoj koncentraciji kalijevih jona. Rezultat je ograničenje ukupnog broja jona u ćeliji. Ako membranski potencijal padne kada je Na/K pumpa blokirana, na primjer, tijekom anoksije, tada se ravnotežni potencijal za hloridne ione smanjuje, a unutarćelijska koncentracija hloridnih jona raste u skladu s tim. Vraćajući ravnotežu naelektrisanja, joni kalijuma takođe ulaze u ćeliju; povećava se ukupna koncentracija iona u ćeliji, što povećava osmotski tlak; ovo tjera vodu da uđe u ćeliju. Ćelija nabubri. Vidi se ovaj otok in vivo u uslovima nedostatka energije.
gradijent koncentracije N / A + kao pokretačka snaga za membranski transport . Značaj Na/K–pumpe za ćeliju nije ograničen na stabilizaciju normalnih K+ i Na+ gradijenata preko membrane. Energija pohranjena u gradijentu membrane Na+ , često se koristi za obezbjeđivanje membranskog transporta za druge supstance. Na primjer, na sl. 1.10 prikazuje "symport" Na+ i molekule šećera u ćeliju. Protein za transport membrane prenosi molekul šećera u ćeliju čak i protiv gradijenta koncentracije, dok se Na+ kreće duž gradijenta koncentracije i potencijala, osiguravajući energiju za transport šećera. Takav transport šećera u potpunosti zavisi od postojanja visokog gradijenta Na+ ; ako je intracelularna koncentracija Na+ značajno se povećava, tada prestaje transport šećera. Za razne c Akharov, postoje različiti sistemi simbola. Transport aminokiselina u kavezu sličnom transportu c akharov prikazan na sl. 1.10; takođe je obezbeđen gradijentom Na+ , postoji najmanje pet različitih sistema simbola, od kojih je svaki specijalizovan za jednu grupu srodnih aminokiselina.
Pored sistema simbola, postoje i sistemi "antiport". Jedan od njih, na primer, prenosi jedan jon kalcijuma iz ćelije u jednom ciklusu u zamenu za tri ulazna jona natrijuma (slika 1.10). Energija za transport Ca 2+ nastaje zbog ulaska tri natrijeva jona duž gradijenta koncentracije i potencijala. Ova energija je dovoljna (u stanju mirovanja) za održavanje visokog gradijenta jona kalcijuma (od manje od 10-7 mol/l unutar ćelije do oko 2 mmol/l izvan ćelije).
Endo- i egzocitoza . Za određene supstance koje ulaze u ćeliju ili se moraju ukloniti

Rice. 1.10.Proteini ugrađeni u lipidni dvosloj membrane posreduju u uvozu glukoze i Na+ u ćeliju, kao i Ca2+/Na+ -antiport, u kojem je pokretačka sila gradijent Na+ na ćelijskoj membrani
od njega nema transportnih kanala; takve supstance uključuju, na primjer, proteine i kolesterol. Mogu proći kroz plazma membranu u vezikule ili mjehurići, endo- i egzocitozom. Na sl. 1.11 prikazuje glavne mehanizme ovih procesa. Tokom egzocitoze, određene organele (vidi dolje) formiraju vezikule ispunjene supstancom koju treba ukloniti iz ćelije, kao što su hormoni ili ekstracelularni enzimi. Kada takve vezikule dođu do plazma membrane, njihova lipidna membrana se stapa s njom, omogućavajući tako sadržaju da pobjegne u vanjsko okruženje. U suprotnom procesu, endocitozi, plazma membrana invaginira, formirajući jamu, koja se zatim produbljuje i zatvara, formirajući unutarćelijsku vezikulu ispunjenu ekstracelularnom tekućinom i nekim makromolekulama. Da bi se osigurala ova membranska fuzija i zatvaranje vezikula, kontraktilni elementi citoskeleta djeluju u sprezi sa samim membranama (vidi dolje). Endocitoza ne uključuje uvijek jednostavno hvatanje ekstracelularnog medija u ćeliju. Ćelijska membrana sadrži, često organizovane u specijalizovane grupe, specifične receptore za makromolekule kao što su insulin ili antigeni. Nakon što se ovi makromolekuli vežu za svoje receptore, dolazi do endocitoze u membranskoj regiji koja okružuje receptor, a makromolekul se selektivno transportuje u ćeliju (slika 1.12, B).
Endo- i egzocitoza se javlja kontinuirano u ćelijama. Količina membranskog materijala koji cirkuliše je značajna; u roku od 1 sata, makrofag apsorbira u obliku vezikula dvostruku površinu svoje citoplazmatske membrane. U većini ćelija promet membranskog materijala nije toliko intenzivan, ali bi ipak trebao biti značajan.

Rice. 1.11.egzocitoza i endocitoza. gore: intracelularna vezikula se spaja sa lipidnim dvoslojem plazma membrane i otvara se u ekstracelularni prostor. Ovaj proces se naziva egzocitoza. Na dnu: plazma membrana invaginira na malom području i spaja vezikulu ispunjenu ekstracelularnim materijalom. Ovaj proces se naziva endocitoza.
1.3. Transport supstanci unutar ćelije
Endo- i egzocitoza nisu samo procesi transporta tvari kroz ćelijsku membranu, već i procesi membranske izmjene - strukturnih komponenti same ćelije. Predmet razmatranja u ovom odeljku su drugi slični transportni procesi u ćeliji i njenim organelama.

Rice. 1.12. A-B. Shema procesa, uključujući egzo- i endocitozu. ALI. Protein sintetizovan u granularnom endoplazmatskom retikulumu transportuje se preko Golgijevog aparata do plazma membrane, gde se izlučuje egzocitozom. B. Holesterol vezan za čestice LDL (lipoproteina niske gustine) vezuje se za plazma membranu, izaziva stvaranje endocitnih vezikula u ovom dijelu membrane i transportuje se do lizozoma, gdje se oslobađa. IN. Ekstracelularni materijal uhvaćen tokom endocitoze (na slici desno), transportuje se kroz ćeliju u vezikulama ili vezikulama i oslobađa se egzocitozom (na slici lijevo)
Difuzija . Naravno, u citosolu, razlika koncentracije se eliminiše difuzijom; isto važi i za tečnosti sadržane u organelama. Zbog visoke koncentracije otopljenog proteina, difuzija je ovdje mnogo sporija nego u vodi. Lipidne membrane – oko ćelije i unutar organela – su dvodimenzionalne tečnosti u kojima se javlja difuzija. Lipidi u dvosloju membrane difundiraju unutar vlastitog sloja, rijetko prelazeći s jednog na drugi. Proteini uronjeni u njih su također prilično pokretni; rotiraju oko ose okomite na membranu ili difundiraju bočno sa vrlo različitim konstantama difuzije, 2-10 000 puta sporije od fosfolipida. Dakle, ako se neki proteini kreću slobodno u lipidnom sloju i istom brzinom kao i sami lipidni molekuli, onda su drugi usidreni, tj. prilično snažno povezan sa citoskeletom. U membrani postoje "trajni" agregati specifičnih proteina, kao što su pre- i postsinaptičke strukture nervnih ćelija. Proteini koji se slobodno kreću mogu se demonstrirati njihovim vezanjem za fluorescentne boje, koje se induciraju da svijetle kratkim osvjetljavanjem male površine membrane kratkim bljeskovima. Takvi eksperimenti pokazuju da se za manje od 1 minute proteini vezani za boju ravnomjerno raspoređuju po membrani na udaljenosti do 10 μm.
Aktivni transport u membranama organela .
Procesi aktivnog transporta, koji igraju vitalnu ulogu u funkcionisanju plazma membrane, odvijaju se i unutar ćelije, u membranama organela. Specifični sadržaji različitih organela nastaju dijelom unutarnjom sintezom, a dijelom aktivnim transportom iz citosola. Jedan primjer potonjeg je Ca 2+ pumpa spomenuta gore u sarkoplazmatskom retikulumu mišićnih stanica. Posebno je interesantno da u slučaju sinteze ATP-a u mitohondrijima, suprotan princip važi za ono što se dešava u ATPaznim pumpama plazma membrane (slika 1.6). Tokom sinteze ATP-a, oksidativni metabolizam dovodi do stvaranja strmog gradijenta H+ na unutrašnjim membranama. Ovaj gradijent je pokretačka snaga za proces koji je obrnut ciklusu pumpanja aktivnog molekularnog transporta: H+ ioni se kreću kroz membranu duž gradijenta, a energija koja se oslobađa kao rezultat toga osigurava sintezu ATP-a iz ADP-a i fosfata. . Rezultirajući ATP, zauzvrat, daje energiju ćeliji, uključujući i za aktivni transport.
Transport u vezikulama . Ćelija ima veliki broj organela i povezanih vezikula (slika 1.1). Ove organele, a posebno vezikule, su u stalnom pokretu, prenoseći svoj sadržaj do drugih organela ili do plazma membrane. Vezikule također mogu migrirati sa ćelijske membrane u organele, kao u endocitozi.
Proces lučenje proteina prikazano na sl. 1.12 ALI. Protein se sintetiše u blizini ćelijskog jezgra na ribosomima povezanim sa endoplazmatskim retikulumom (tzv. granularni, ili grubi, endoplazmatski retikulum); Jednom u endoplazmatski retikulum, protein se pakuje u transportne vezikule, koje se odvajaju od organele i migriraju u Golgijev aparat. Ovdje se spajaju sa rezervoarima Golgijevog aparata, gdje se protein modificira (tj. pretvara u glikoprotein). Na krajevima cisterni, vezikule se ponovo odvajaju. Sekretorne vezikule koje nose modifikovani protein kreću se prema plazma membrani i oslobađaju svoj sadržaj egzocitozom.
Još jedan primjer transportnog puta u ćeliji prikazan je na Sl. 1.12, B; je uzimanje holesterola od strane ćelije. Holesterol koji se transportuje u krvi uglavnom je povezan s proteinima, kao što su čestice "lipoprotein niske gustine"(LNP). Ove čestice se vežu za specifična mjesta LDL receptora na membrani gdje se javlja endocitoza i LDL se transportuje u ćeliju u "obloženim" vezikulama. Ove vezikule se spajaju i formiraju endozome i gube svoj 'zvon' tokom ovog procesa. Endosomi se zauzvrat spajaju s primarnim lizosomima koji sadrže pretežno hidrolitičke enzime i formiraju sekundarne, veće lizozome. U njima se kolesterol oslobađa iz LDL čestica i difundira u citosol, gdje postaje dostupan, na primjer, za sintezu lipidnih membrana. Od endozoma se odvajaju i vezikule koje ne sadrže LDL, koji se na poseban način kreću do plazma membrane i spajaju se s njom, vraćajući materijal membrane i, vjerovatno, LDL receptore. Od trenutka kada se LDL čestica veže za membranu, prođe 10-15 minuta dok se holesterol ne oslobodi iz sekundarnog lizosoma. Poremećaji u vezivanju i apsorpciji LDL-a, odnosno u opskrbi ćelije holesterolom, igraju odlučujuću ulogu u nastanku ozbiljne i rasprostranjene bolesti, ateroskleroze („otvrdnuće“ arterija).
Postoji mnogo drugih transportnih ruta sličnih onima prikazanim na sl. 1.11 i 1.12,A, uz pomoć kojih se specifični vezikuli kreću u ćeliji. Ne zna se tačno kako se kreću, ali su elementi citoskeleta vjerovatno uključeni u ovaj proces. Vezikule mogu kliziti duž mikrotubula, u kom slučaju se čini da energiju za kretanje obezbjeđuje protein povezan sa vezikulama, ATPaza (vidi dolje). Ostaje potpuno neshvatljivo koliko različitih vezikula, krećući se jedna za drugom u svim smjerovima, stigne do svog odredišta. Očigledno ih treba „označiti“ na način da ih transportni sistem prepozna i pretvori u svrsishodno kretanje.
Transport formiranjem i uništavanjem organela . Do sada smo endo- i egzocitozu razmatrali kao procese transporta sadržaja vezikula. Postoji još jedan aspekt ovih procesa, koji se sastoji u tome da usmjereno uklanjanje plazma membrane u jednom području ćelijske površine endocitozom i, naprotiv, njeno dodavanje u drugo putem egzocitoze, pomjera značajne dijelove membrana (slika 1.12.E), dajući ćeliji priliku, na primjer, da formira izraslinu ili da se pomakne.
Slična preuređivanja su tipična i za citoskelet, posebno za mikrofilamente i mikrotubule (slika 1.1). Mikrofilamenti sastoji se prvenstveno od F-aktin protein koji je sposoban da se sklapa u fibrozne snopove kao rezultat polimerizacije monomera iz citosola. Snopići su polarizirani, odnosno često rastu samo s jednog kraja, akumulirajući nove molekule aktina, dok je drugi kraj inertan ili se ovdje rastavlja. Zbog ovog polariziranog rasta, mikrofilamenti se efikasno kreću i struktura njihove mreže se može promijeniti. Prijelaz aktina iz depolimeriziranog stanja (sol) u organizirano stanje (gel) može se dogoditi vrlo brzo pod utjecajem drugih proteina ili promjena u koncentraciji jona (vidi dolje). Postoje i proteini koji uzrokuju raspad aktinskih filamenata na kratke fragmente. Tanke izrasline mnogih ćelija – filopodije – sadrže centralni snop aktina (slika 1.1), a različita kretanja filopodija su verovatno posledica aktinskih prelaza: polimerizacija – depolimerizacija.
mikrotubuletakođer se često podvrgavaju sličnim pokretima. Mehanizam ovih kretanja je sličan - polimerizacija tubulina iz citosola na način da jedan od krajeva mikrotubula raste, dok se drugi ili ne mijenja, ili se tamo rastavlja. Dakle, mikrotubul, odgovarajućim dodavanjem ili uklanjanjem materijala, može da se kreće kroz citosol.
Aktivni pokreti citoskeleta . Promjene u strukturama citoskeleta mogu nastati kao rezultat i aktivnih pokreta i preuređivanja opisanih gore. U mnogim slučajevima, kretanje mikrotubula i aktinskih filamenata pokreću kontraktilni proteini koji vežu filamente ili tubule i mogu ih pomicati jedni u odnosu na druge. Vjeverice miozina i dineina prisutan u citosolu svih ćelija u relativno visokim koncentracijama; oni su elementi koji pretvaraju energiju u kretanje u specijalizovanim ćelijama (mišići) i organele (cilije). U mišićnim ćelijama miozin formira debele filamente orijentisane paralelno sa aktinskim filamentima. Molekul miozina se svojom "glavom" veže za aktinski filament i, koristeći energiju ATP, istiskuje miozin duž molekule aktina. Miozin se tada odvaja od aktina. Skup mnogih takvih ciklusa spajanja-odspajanja vodi do makroskopskog kontrakcija mišićnih vlakana(Pogl. 4). Dinein igra sličnu ulogu u kretanju mikrotubula tokom operacije cilija (slika 1.1). U citoplazmi nespecijaliziranih stanica miozin i dinein ne formiraju pravilna vlakna, već u većini slučajeva male grupe molekula. Čak iu obliku tako malih agregata, oni su u stanju pomicati aktinske filamente ili mikrotubule. Rice. 1.13 ilustruje ovaj proces kada su suprotno polarizovani molekuli miozina takođe vezani za dva aktinska filamenta polarizovana u suprotnim smerovima. Glavne grupe miozina se savijaju prema repu molekula, trošeći ATP, a dva aktinska filamenta se pomiču u suprotnom smjeru, nakon čega se miozin odvaja od njih. Pokreti ove vrste, tokom kojih se energija ATP-a pretvara u mehanički rad, mogu promijeniti oblik citoskeleta, a samim tim i ćelija, kao i osigurati transport organela povezanih sa citoskeletom.
Procesi intracelularnog transporta mogu se najjasnije pokazati na aksonu nervne ćelije. transport aksona ovdje se detaljno raspravlja kako bi se ilustrovali događaji koji se vjerovatno događaju na sličan način u većini ćelija. Akson koji ima samo nekoliko mikrona u prečniku može biti dugačak jedan metar ili više, a potrebne su godine da se proteini difuzijom kreću od jezgra do distalnog kraja aksona. Odavno je poznato da kada se bilo koji dio aksona steže, proksimalni dio aksona se širi. Izgleda da je centrifugalni tok blokiran u aksonu. Takve tok-brz transport aksona može može se pokazati kretanjem radioaktivnih markera, kao u eksperimentu prikazanom na Sl. 1.14. Radioaktivno obilježen leucin ubrizgan je u ganglij dorzalnog korijena, a zatim je od 2. do 10. sata mjerena radioaktivnost u išijadičnom živcu na udaljenosti od 166 mm od tijela neurona. Tokom 10 sati, vrh radioaktivnosti na mjestu injekcije nije se značajno promijenio. Ali talas radioaktivnosti se širio duž aksona konstantnom brzinom od oko 34 mm u 2 sata, ili 410 mm/dan. Pokazalo se da se u svim neuronima homoiotermnih životinja brzi transport aksona odvija istom brzinom, te da nema primjetnih razlika između tankih, nemijeliniziranih vlakana i najdebljih aksona, kao ni između motornih i senzornih vlakana. Tip radioaktivnog markera takođe ne utiče na brzinu brzog aksonalnog transporta; Kao markeri mogu poslužiti razne radioaktivne supstance.

Rice. 1.13.Nemišićni miozinski kompleks s određenom orijentacijom može se vezati za aktinske filamente različitog polariteta i, koristeći energiju ATP-a, pomjeriti ih jedan u odnosu na drugi.
molekule, kao što su različite aminokiseline, koje su ugrađene u tjelesne proteine neurona. Ako analiziramo periferni dio živca da bismo utvrdili prirodu nositelja radioaktivnosti koja se ovdje prenosi, onda se takvi nosioci nalaze uglavnom u proteinskoj frakciji, ali iu sastavu medijatora i slobodnih aminokiselina. Znajući da su svojstva ovih supstanci različita, a posebno veličine njihovih molekula različite, konstantnu brzinu transporta možemo objasniti samo transportnim mehanizmom koji je zajednički za sve njih.
Gore opisano brz transport aksona je anterogradni tj. usmjerena dalje od tijela ćelije. Pokazalo se da se neke supstance kreću sa periferije u telo ćelije uz pomoć retrogradni transport. Na primjer, acetilholinesteraza se transportuje u ovom smjeru dvostruko manjom brzinom od brzine brzog aksonalnog transporta. Marker koji se često koristi u neuroanatomiji - peroksidaza hrena - također se kreće retrogradno. Retrogradni transport vjerovatno igra važnu ulogu u regulaciji sinteze proteina u ćelijskom tijelu. Nekoliko dana nakon transekcije aksona, u ćelijskom tijelu se opaža hromatoliza, što ukazuje na kršenje sinteze proteina. Vrijeme potrebno za hromatolizu korelira s trajanjem retrogradnog transporta od mjesta transekcije aksona do tijela ćelije. Takav rezultat također sugerira objašnjenje za ovo kršenje - poremećen je prijenos s periferije "signalne supstance" koja regulira sintezu proteina. Očigledno, glavna "vozila" se koriste za brzi aksonal
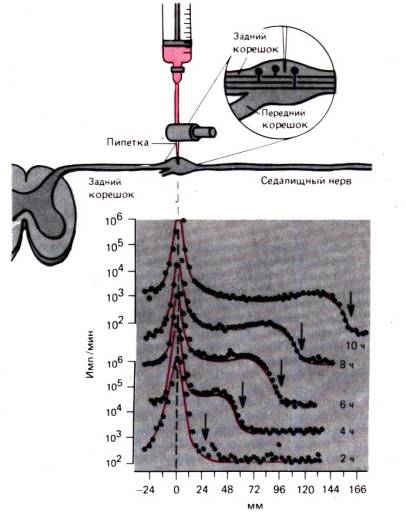
Rice. 1.14.Eksperiment koji demonstrira brz transport aksona u senzornim vlaknima išijadičnog živca mačke. Leucin obeležen tricijumom se ubrizgava u dorzalni ganglij korena, a radioaktivnost u gangliju i senzornim vlaknima se meri 2, 4, 6, 8 i 10 sati nakon injekcije. (donji deo slike). By apscisa odgađa se udaljenost od ganglija do dijelova išijadičnog živca, gdje se vrši mjerenje. Na y-osi, samo za gornju i donju krivu, radioaktivnost (imp./min) je ucrtana u logaritamskoj skali. "Talas" povećane radioaktivnosti (strelice) kreće se brzinom od 410 mm/dan (do )
transport su vezikule (vezikule) i organele, kao što su mitohondrije koje sadrže supstance koje se transportuju. Pokret najvećih vezikula ili mitohondrija može se posmatrati pomoću mikroskopa in vivo . Takve čestice prave kratke, brze pokrete u jednom smjeru, zaustavljaju se, često se pomiču malo unatrag ili u stranu, ponovo se zaustavljaju, a zatim jure u glavnom smjeru. 410 mm/dan odgovara prosječnoj anterogradnoj brzini od približno 5 μm/s; brzina svakog pojedinačnog pokreta bi stoga trebala biti mnogo veća, a ako uzmemo u obzir veličinu organela, filamenata i mikrotubula, onda su ti pokreti zaista vrlo brzi. Brzi transport aksona zahtijeva značajnu koncentraciju ATP-a. Otrovi kao što je kolhicin koji uništavaju mikrotubule također blokiraju brzi transport aksona. Iz ovoga proizilazi da se u procesu transporta koji razmatramo, vezikule i organele kreću duž mikrotubula i aktinskih filamenata; ovo kretanje osiguravaju mali agregati molekula dineina i miozina koji djeluju kao što je prikazano na sl. 1.13, koristeći energiju ATP-a.
Brzi transport aksona također može biti uključen patoloških procesa. Neki neurotropni virusi (na primjer, herpes ili polio virusi) prodiru u akson na periferiji i kreću se retrogradnim transportom do tijela neurona, gdje se umnožavaju i ispoljavaju svoje toksično djelovanje. Tetanus toksin, protein proizveden od strane bakterija koji ulazi u tijelo kroz lezije na koži, preuzimaju ga nervni završeci i transportuju do tijela neurona, gdje izaziva karakteristične grčeve mišića. Poznati su slučajevi toksičnih efekata na sam transport aksona, na primer, izlaganje industrijskom rastvaraču akrilamidu. Osim toga, vjeruje se da patogeneza beri-beri beri-beri i alkoholne polineuropatije uključuje kršenje brzog aksonalnog transporta.
Pored brzog transporta aksona u ćeliji postoji i prilično intenzivan spor transport aksona. Tubulin se kreće duž aksona brzinom od oko 1 mm/dan, dok se aktin kreće brže, do 5 mm/dan. Sa ovim komponentama citoskeleta migriraju i drugi proteini; na primjer, čini se da su enzimi povezani s aktinom ili tubulinom. Brzine kretanja tubulina i aktina su otprilike u skladu sa stopama rasta pronađenim za mehanizam koji je ranije opisan kada su molekuli ugrađeni u aktivni kraj mikrotubula ili mikrofilamenta. Stoga ovaj mehanizam može biti u osnovi sporog aksonalnog transporta. Brzina sporog transporta aksona također približno odgovara brzini rasta aksona, što, očigledno, ukazuje na ograničenja koja struktura citoskeleta nameće drugom procesu.
Završavajući ovaj dio, treba naglasiti da ćelije nikako nisu statične strukture, kako izgledaju, na primjer, na elektronskim mikroskopskim fotografijama. plazma membrana a posebno organele su u stalnom brzom kretanju i stalnom restrukturiranju; to je jedini razlog zašto su u stanju da funkcionišu. Dalje, to nisu jednostavne komore u kojima se odvijaju hemijske reakcije, već visoko organizovani konglomerati membrana i vlakana, u kojoj se reakcije odvijaju u optimalno organizovanom nizu.
1.4. Regulacija ćelijskih funkcija
Održavanje individualne ćelije kao funkcionalne jedinice u velikoj meri je regulisano jezgrom; proučavanje takvih regulatornih mehanizama je predmet ćelijske biologije i biohemije. Istovremeno, ćelije moraju da modifikuju svoje funkcije u skladu sa uslovima sredine i potrebama drugih ćelija tela, odnosno služe kao objekti funkcionalne regulacije. U nastavku ćemo ukratko razmotriti kako ovi regulatorni utjecaji djeluju na plazma membranu i kako dopiru do intracelularnih organela.
Regulatorni efekti na ćelijsku membranu
Potencijal membrane . U mnogim slučajevima, regulacija ćelijskih funkcija se provodi promjenom membranskog potencijala. Lokalne promene potencijala su moguće kada: 1) kroz membranu teče struja iz susedne ćelije ili koju generiše druga ćelija; 2) mijenja se ekstracelularna koncentracija jona (često [K + ] van ); 3) membranski jonski kanali otvoreni. Promjene u membranskom potencijalu mogu utjecati na konformaciju membranskih proteina, uzrokujući, posebno, otvaranje ili zatvaranje kanala. Kao što je gore opisano, funkcioniranje nekih membranskih pumpi ovisi o potencijalu membrane. Nervne ćelije su specijalizovane da percipiraju promene u membranskom potencijalu kao informaciju koja se mora obraditi i preneti (vidi Poglavlje 2).
ekstracelularne regulatorne supstance . Najvažniji regulatorni mehanizam koji uključuje ekstracelularne supstance je njihova interakcija sa specifičnim receptorima na plazma membrani ili unutar ćelije. Ove supstance uključuju sinaptičke medijatore koji prenose informacije između nervnih ćelija, lokalnih agenasa i supstanci koje cirkulišu u krvi i dospevaju u sve ćelije u telu, kao što su hormoni i antigeni. sinaptičkih neurotransmitera su male molekule koje se oslobađaju iz nervnih završetaka u sinapsi;
kada dođu do plazma membrane susjedne postsinaptičke ćelije, pokreću električne signale ili druge regulatorne mehanizme. Ovo pitanje je detaljno razmotreno u Pogl. 3.
Lokalni hemijski agensi često luče specijalizovane ćelije. Oni slobodno difundiraju u ekstracelularnom prostoru, ali njihovo djelovanje je ograničeno na malu grupu stanica zbog brzog uništavanja ovih supstanci, bilo spontano ili djelovanjem enzima. Jedan primjer oslobađanja takvih agenasa je oslobađanje histamin mastociti nakon ozljede ili imunološkog odgovora. Histamin izaziva relaksaciju vaskularnih glatkih mišićnih ćelija, povećava permeabilnost vaskularnog endotela i stimuliše senzorne nervne završetke koji posreduju u osećaju svraba. Druge lokalne hemijske agense luče mnoge druge ćelije. Tipični lokalni agenti su prostaglandini,čine grupu od otprilike 20 derivata masnih kiselina. Oslobađaju se kontinuirano iz široko rasprostranjenih stanica, ali djeluju samo lokalno, jer se brzo uništavaju membranskim fosfolipazama. Različiti prostaglandini imaju širok spektar djelovanja: mogu pokrenuti kontrakciju glatkih mišićnih stanica, uzrokovati agregaciju krvnih pločica (trombocita) ili inhibirati razvoj žutog tijela u jajnicima.
Drugi lokalni agenti služe faktori rasta. Najpoznatiji faktor rasta nerava (NGF) za simpatičke neurone, koji je neophodan za rast i opstanak ovih neurona tokom razvoja in vivo ili u ćelijskoj kulturi. Očigledno, ciljne ćelije za ovu klasu neurona luče NGF i na taj način obezbeđuju ispravnu inervaciju. Prilikom formiranja organa, stanice često moraju "naći put" do ciljnih stanica, koje se mogu nalaziti na znatnoj udaljenosti. Shodno tome, mora postojati mnogo specijalizovanih faktora rasta kao što je NGF.
Hormoni i antigeni prenosi krv do svih ćelija. Antigeni izazivaju imuni odgovor ćelija koje nose specifična antitijela. Međutim, antigeni su, po pravilu, strane supstance koje se ne formiraju u reagujućem organizmu (za više detalja videti Poglavlje 18). Neki hormoni, kao što su insulin ili tiroksin, utiču na širok spektar tipova ćelija, dok drugi, kao što su polni hormoni, utiču samo na određene tipove ćelija. Hormoni su ili peptidi, čije djelovanje je pokrenuto njihovim vezanjem za receptor na ćelijskoj membrani, ili steroidi i tiroksin, koji difundiraju kroz lipidnu membranu i vezuju se za intracelularne receptore. Steroidni hormoni se vežu za nuklearni hromatin, što rezultira transkripcijom određenih gena. Proteini koji nastaju kao rezultat uzrokuju promjenu staničnih funkcija, što je specifično djelovanje hormona. Pitanja vezana za oslobađanje i djelovanje hormona detaljno su razmotrena u Pogl. 17.
Intracelularna komunikacija koja uključuje sekundarne glasnike
Gore opisane regulatorne funkcije uključuju efekte na ćelijsku membranu. Informacije koje prima ćelijska membrana često moraju izazvati reakciju u organelama i do njih se prenose različitim supstancama poznatim kao drugi glasnici (za razliku od prvih, koji u ćeliju dolaze iz vanjskih izvora). Studija o drugim medijatorima se ubrzano razvija i nema garancije da će sadašnji nivo razumijevanja problema biti dovoljno potpun. Ovdje ćemo se dotaknuti tri dobro proučavana medijatora: Ca 2+, cAMP i inozitol trifosfat.
Kalcijum.Najjednostavniji intracelularni medijator je ion Ca 2+. Njegova slobodna koncentracija u stanici u mirovanju je vrlo niska i iznosi 10 -8 -10 -7 mol/l. Može ući u ćeliju kroz specifične membranske kanale kada su otvoreni, na primjer, kada se promijeni membranski potencijal (vidi Poglavlje 2). Rezultirajuće povećanje Ca 2+ pokreće važne reakcije u ćeliji, kao što je kontrakcija miofibrila, koja je osnova kontrakcije mišića (vidi Poglavlje 4), ili oslobađanje vezikula koji sadrže neurotransmiter iz nervnih završetaka (vidi Poglavlje 3). . Obje reakcije zahtijevaju koncentraciju Ca 2+ od približno 10-5 mol/l. Ca 2+, koji ima regulatorno dejstvo, takođe se može osloboditi iz intracelularnih depoa, kao što je endoplazmatski retikulum. Oslobađanje Ca 2+ iz depoa zahteva učešće drugih posrednika (vidi, na primer, sliku 1.16).
Ciklični adenozin monofosfat, cAMP. Nedavno je dokazano da je ciklički adenozin monofosfat (cAMP), derivat glavnog izvora energije u tijelu, ATP, važan drugi glasnik. Složeni lanac reakcija prikazan na sl. 1.15, počinje od receptora Rs na vanjskoj površini plazma membrane, koja može poslužiti kao specifično mjesto vezivanja za različite medijatore i hormone. Nakon vezivanja za specifičan "stimulirajući" molekul Rs mijenja svoju konformaciju; ove promjene utiču na protein Gs na unutrašnjoj površini membrane na takav način da postaje moguće aktivirati potonju intracelularnim gvanozin trifosfatom (GTP). aktivirani protein Gs , zauzvrat, stimulira enzim na unutrašnjoj površini membrane, adenilat ciklazu (AC), koji katalizira stvaranje cAMP iz ATP-a. CAMP rastvorljiv u vodi i posrednik je koji prenosi efekat

Rice. 1.15.Lanac reakcija koji uključuje intracelularni medijator cAMP (ciklički adenozin monofosfat). Ekscitatorni ili inhibitorni vanjski signali aktiviraju membranske receptore R s ili Ri . Ovi receptori regulišu proces vezivanja G -proteini sa intracelularnim GTP (gvanozin trifosfat), čime stimulišu ili inhibiraju intracelularnu adenilat ciklazu (AC). Enzim koji pojačava AC pretvara adenozin trifosfat (ATP) u cAMP, koji se zatim cijepa na AMP uz pomoć fosfodiestera. Slobodni cAMP difundira u ćeliju i aktivira adenilat kinazu (A-kinazu), oslobađajući njenu katalitičku podjedinicu C, koja katalizuje fosforilaciju intracelularnih proteina, tj. formira konačni efekat ekstracelularnog stimulusa. Shema također prikazuje farmakološke lijekove i toksine koji pokreću (+) ili inhibiraju (-) neke reakcije (ali s promjenama)
stimulacija ekstracelularnog receptora Rs na unutrašnje strukture ćelije.
Paralelno sa stimulativnim lancem reakcija koje uključuju Rs moguće vezivanje inhibitornih medijatora i hormona za odgovarajući receptor R i koji opet putem GTP-aktiviranog proteina G , inhibira AC i time proizvodnju cAMP. Difundirajući u ćeliju, cAMP reaguje sa adenilat kinazom (A-kinazom); Ovo oslobađa C podjedinicu, koja katalizira fosforilaciju proteina. Ova fosforilacija pretvara proteine u njihov aktivni oblik i sada oni mogu vršiti svoje specifično regulatorno djelovanje (na primjer, uzrokovati razgradnju glikogena). Ovaj složeni regulatorni sistem je izuzetno efikasan, jer je krajnji rezultat fosforilacija mnogih proteina, odnosno, regulatorni signal prolazi kroz lanac sa velikim faktorom pojačanja. Vanjski posrednici koji se vezuju za receptore R s i R i specifične za svaku od njih su izuzetno raznolike. Adrenalin, petljanje sa R s ili R i učestvuje u regulaciji metabolizma lipida i glikogena, kao iu pojačanoj kontrakciji srčanog mišića i drugim reakcijama (vidi Poglavlje 19). hormon koji stimuliše štitnjaču, aktiviranje Rs , stimuliše lučenje hormona tiroksina od strane štitne žlezde, a prostaglandin I inhibira agregaciju trombocita. Inhibicijski efekti, uključujući adrenalin, posredovani R i izraženo u usporavanju lipolize. Na ovaj način, cAMP sistem je multifunkcionalni unutarćelijski regulatorni sistem, koji se može precizno kontrolisati ekstracelularnim stimulativnim i inhibitornim signalnim agensima.
Inozitol fosfat "IF h ". Intracelularni sistem drugog medijatora, inozitol fosfata, tek je nedavno otkriven (slika 1.16). U ovom slučaju ne postoji inhibitorni put, ali postoji sličnost sa cAMP sistemom, u kojem se efekat stimulacije R receptora prenosi na GTP-aktivirani G-protein na unutrašnjoj površini membrane. U sljedećoj fazi, uobičajeni membranski lipidni fosfatidilinozitol (PI), koji je prethodno primio dvije dodatne fosfatne grupe, pretvara se u PI-difosfat (FIF 2), koji se aktiviranom fosfodiesterazom (PDE) cijepa u inozitol trifosfat(IFz) i lipida diacilglicerol(DAG). Inozitol trifosfat je vodotopivi sekundarni glasnik koji difundira u citosol. Djeluje prvenstveno oslobađanjem Ca 2+ iz endoplazmatskog retikuluma. Ca 2+ zauzvrat djeluje kao posrednik kao što je gore opisano; na primjer, aktivira Ca2+-zavisnu fosfokinazu, koja fosforiliše enzime. Lipidna podjedinica DAG-a (slika 1.16) takođe nosi signal, difundirajući u lipidnoj fazi plazma membrane do C-kinaze koja se nalazi na njenoj unutrašnjoj površini, a koja se aktivira uz učešće fosfatidilserina kao kofaktora. Tada C-kinaza pokreće fosforilaciju proteina, pretvarajući ih u aktivni oblik.
Intracelularni sistem drugog intermedijarnog interferona 3 se takođe može kontrolisati raznim eksternim medijatorima i hormonima, uključujući acetilholin, serotonin, vazopresin i hormon koji stimuliše štitnjaču; kao i cAMP sistem, karakteriše ga niz intracelularnih efekata. Moguće je da se ovaj sistem takođe aktivira svetlošću u vizuelnom receptoru oka i da igra centralnu ulogu u fototransdukciji (videti Poglavlje 11). Po prvi put u individualnom razvoju organizma, receptor IGF sistema se aktivira spermatozoidima, usled čega IGF učestvuje u regulatornim reakcijama koje prate oplodnju jajne ćelije.
cAMP i IFz-DAG sistemi su veoma efikasni biološki pojačivači. Oni su

Rice. 1.16.Lanac reakcija koji uključuje intracelularni intermedijarni interferon (inozitol trifosfat). Kao iu cAMP sistemu, ekstracelularni signal je posredovan preko proteinaG, koji u ovom slučaju aktivira fosfodiesterazu (PDE). Ovaj enzim razgrađuje fosfatidilinozin difosfat (FIF) 2 ) u plazma membrani prije IF h i diacilglicerol (DAG); IF h difunduje u citoplazmu. Ovdje on izaziva oslobađanje Ca 2+ iz endoplazmatskog retikuluma; povećanje koncentracije Ca 2+ u citoplazmi ([Ca 2+ ] i ) aktivira protein kinazu koja fosforilira i stoga aktivira enzime. Drugi proizvod, DAG, ostaje u membrani i aktivira protein kinazu C (kofaktor-fosfatidilserin, PS). Protein kinaza C također fosforilira enzime koji posreduju u specifičnim akcijama povezanim sa stimulacijom vanjskog receptora. R . Grane lanca reakcija koje uključuju IF h i DAG se mogu nezavisno aktivirati jonomicinom i forbol esterom, respektivno (modifikovani)
pretvaraju reakciju između medijatora i receptora vanjske membrane u fosforilaciju mnogih intracelularnih proteina, koji potom mogu utjecati na različite ćelijske funkcije. Jedan od bitnih aspekata problema je da, koliko danas znamo, postoje samo ova dva blisko povezana regulatorna sistema ovog tipa, koje koriste brojni eksterni posrednici za regulaciju različitih unutarćelijskih procesa. Istovremeno, ovi regulatorni sistemi, uključujući Ca 2+, usko su u interakciji jedni s drugima, što im omogućava finu regulaciju ćelijskih funkcija.
Tutorijali i vodiči
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts TO., Watson J.D.Molekularna biologija ćelije, Njujork i London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(ur.). Biologija. Berlin, Hajdelberg, Njujork, Springer, 1983.
3. hille IN. Jonski kanali ekscitabilnih membrana. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W. Marki H., Ziegler H.(ur.). Biophysik. Berlin, Hajdelberg, Njujork, Springer, 1984.
5. Jungermann TO., Mahler H.Biohemija. Berlin, Hajdelberg, Njujork, Springer, 1980.
6. Kandel E.R., Schwartz-J.H.,(ur.). Principi neuronauke, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., SchmidtW.Anatomical des Menschen. Berlin, Hajdelberg, Njujork, Tokio, Springer, 1983.
Originalni članci i recenzije
8. Berridge M.J. Molekularna osnova komunikacije unutar ćelije, Sci. Amer 253 124 134 (1985).
9. Berridge M. J., Irvine R. F. Inozitol trifosfat, novi drugi glasnik u transdukciji ćelijskog signala. Nature, 312, 315 321 (1984).
10. Bretscher M.S. Molekuli ćelijske membrane, Sci. Amer., 253, 124–134 (1985).
11. Daut J.Živa ćelija kao mašina za pretvaranje energije. Minimalni model metabolizma miokarda, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz IN. Utjecaj jona natrijuma na električnu aktivnost gigantskog aksona lignje. J Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Aktivni transport kationa u džinovskim aksonima iz Sepia i Loligo, J Physiol. (Lond.), 128, 28–42 (1955).
14. Duži P. Jonski kanali sa konformacionim podstanjima, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Egzoplazmatski transport u normalnim i patološkim sistemima. U: Fiziologija i patologija aksona, S.G. Waxman, Ed. Njujork, Raven Press, 1978.
Pozivamo vas da se upoznate sa materijalima i.
: celulozna membrana, membrana, citoplazma sa organelama, jezgro, vakuole sa ćelijskim sokom.Prisustvo plastida je glavna karakteristika biljne ćelije.
Funkcije ćelijskog zida- određuje oblik ćelije, štiti od faktora okoline.
plazma membrana- tanak film, koji se sastoji od molekula lipida i proteina u interakciji, omeđuje unutrašnji sadržaj od spoljašnje sredine, obezbeđuje transport vode, mineralnih i organskih materija u ćeliju putem osmoze i aktivnog prenosa, a takođe uklanja otpadne proizvode.
Citoplazma- unutrašnje polutečno okruženje ćelije, u kojem se nalaze jezgro i organele, obezbeđuje veze između njih, učestvuje u glavnim procesima života.
Endoplazmatski retikulum- mreža razgranatih kanala u citoplazmi. Učestvuje u sintezi proteina, lipida i ugljikohidrata, u transportu tvari. Ribosomi - tijela smještena na EPS ili u citoplazmi, sastoje se od RNK i proteina, uključena su u sintezu proteina. EPS i ribozomi su jedan aparat za sintezu i transport proteina.
Mitohondrije-organele odvojene od citoplazme sa dvije membrane. U njima se oksidiraju organske tvari i sintetiziraju se molekule ATP-a uz sudjelovanje enzima. Povećanje površine unutrašnje membrane na kojoj se nalaze enzimi zbog krista. ATP je organska supstanca bogata energijom.
plastidi(hloroplasti, leukoplasti, hromoplasti), njihov sadržaj u ćeliji je glavna karakteristika biljnog organizma. Kloroplasti su plastidi koji sadrže zeleni pigment hlorofil, koji apsorbira svjetlosnu energiju i koristi je za sintezu organskih tvari iz ugljičnog dioksida i vode. Razgraničenje hloroplasta od citoplazme pomoću dvije membrane, brojnih izraslina - grana na unutrašnjoj membrani, u kojoj se nalaze molekule hlorofila i enzimi.
Golgijev kompleks- sistem šupljina ograničenih od citoplazme membranom. Akumulacija proteina, masti i ugljikohidrata u njima. Implementacija sinteze masti i ugljikohidrata na membranama.
Lizozomi- tijela odvojena od citoplazme jednom membranom. Enzimi sadržani u njima ubrzavaju reakciju cijepanja složenih molekula na jednostavne: proteina do aminokiselina, složenih ugljikohidrata do jednostavnih, lipida do glicerola i masnih kiselina, a također uništavaju mrtve dijelove stanice, cijele stanice.
Vakuole- šupljine u citoplazmi ispunjene ćelijskim sokom, mjesto nakupljanja rezervnih hranjivih tvari, štetnih tvari; regulišu sadržaj vode u ćeliji.
Core- glavni dio ćelije, sa vanjske strane prekriven dvomembranom, probušen porama nuklearnog omotača. Supstance ulaze u jezgro i uklanjaju se iz njega kroz pore. Kromosomi su nosioci nasljednih informacija o karakteristikama organizma, glavnim strukturama jezgra, od kojih se svaki sastoji od jednog molekula DNK u kombinaciji s proteinima. Jezgro je mjesto sinteze DNK, i-RNA, r-RNA.

Prisustvo vanjske membrane, citoplazme sa organelama, jezgra sa hromozomima.
Vanjska ili plazma membrana- omeđuje sadržaj ćelije od okoline (ostale ćelije, međućelijska supstanca), sastoji se od molekula lipida i proteina, obezbeđuje komunikaciju između ćelija, transport supstanci u ćeliju (pinocitoza, fagocitoza) i van ćelije.
Citoplazma- unutrašnje polutečno okruženje ćelije, koje obezbeđuje komunikaciju između jezgra i organela koji se nalaze u njoj. Glavni procesi vitalne aktivnosti odvijaju se u citoplazmi.
ćelijske organele:
1) endoplazmatski retikulum (ER)- sistem granajućih tubula, uključenih u sintezu proteina, lipida i ugljenih hidrata, u transportu supstanci u ćeliji;
2) ribozomi- tijela koja sadrže rRNA nalaze se na ER i u citoplazmi, te su uključena u sintezu proteina. EPS i ribozomi su jedan aparat za sintezu i transport proteina;
3) mitohondrije- "elektrane" ćelije, odvojene od citoplazme sa dve membrane. Unutrašnji formira kriste (nabore) koje povećavaju njegovu površinu. Enzimi na kristama ubrzavaju reakcije oksidacije organskih supstanci i sintezu energetski bogatih molekula ATP-a;
4) golgi kompleks- grupa šupljina ograničenih membranom od citoplazme, ispunjenih proteinima, mastima i ugljikohidratima, koji se ili koriste u životnim procesima ili uklanjaju iz ćelije. Membrane kompleksa vrše sintezu masti i ugljikohidrata;
5) lizozomi- tijela ispunjena enzimima ubrzavaju reakcije razgradnje proteina do aminokiselina, lipida do glicerola i masnih kiselina, polisaharida do monosaharida. U lizosomima se uništavaju mrtvi dijelovi ćelije, cijele ćelije i ćelije.
Uključci ćelija- Akumulacije rezervnih nutrijenata: proteina, masti i ugljenih hidrata.
Core- najvažniji deo ćelije. Prekrivena je dvomembranskom membranom s porama kroz koje neke tvari prodiru u jezgro, dok druge ulaze u citoplazmu. Hromozomi su glavne strukture jezgra, nosioci nasljednih informacija o karakteristikama organizma. Prenosi se u procesu diobe matične ćelije na ćerke ćelije, a sa zametnim ćelijama - na ćerke organizme. Jezgro je mjesto sinteze DNK, mRNA, rRNA.
Zadatak:
Objasnite zašto se organele nazivaju specijalizovanim strukturama ćelije?
odgovor: Organele se nazivaju specijalizirane ćelijske strukture, budući da obavljaju strogo određene funkcije, nasljedne informacije se pohranjuju u jezgri, ATP se sintetizira u mitohondrijima, fotosinteza se odvija u kloroplastima itd.
Ako imate pitanja o citologiji, možete zatražiti pomoć od
Treća faza evolucije je pojava ćelije.
Molekuli proteina i nukleinskih kiselina (DNK i RNK) formiraju biološku ćeliju, najmanju jedinicu života. Biološke ćelije su "građevinski blokovi" svih živih organizama i sadrže sve materijalne kodove razvoja.
Naučnici su dugo vremena smatrali da je struktura ćelije izuzetno jednostavna. Sovjetski enciklopedijski rečnik tumači koncept ćelije na sledeći način: "Ćelija je elementarni živi sistem, osnova strukture i života svih životinja i biljaka." Treba napomenuti da izraz "elementaran" ni na koji način ne znači "jednostavan". Naprotiv, ćelija je jedinstvena fraktalna tvorevina Boga, koja zadivljuje svojom složenošću i, istovremeno, izuzetnom koherentnošću rada. svih njegovih elemenata.
Kada smo uz pomoć elektronskog mikroskopa uspjeli pogledati unutra, pokazalo se da je uređaj najjednostavnije ćelije složen i neshvatljiv kao i sam Univerzum. Danas je već utvrđeno da je "ćelija posebna materija Univerzuma, posebna materija Kosmosa". Jedna jedina ćelija sadrži informacije koje se mogu staviti u samo nekoliko desetina hiljada tomova Velike sovjetske enciklopedije. One. ćelija je, između ostalog, ogroman "biorezervoar" informacija.
Autor moderne teorije molekularne evolucije, Manfred Eigen, piše: „Da bi se slučajno formirao proteinski molekul, priroda bi morala izvršiti oko 10130 pokušaja i potrošiti na to toliki broj molekula koji bi bio dovoljan za 1027 Univerzumi.Ako je protein izgrađen inteligentno, odnosno da se valjanost svakog poteza može provjeriti nekim selekcionim mehanizmom, bilo je potrebno samo oko 2000 pokušaja.Dolazimo do paradoksalnog zaključka: program za izgradnju "primitivne žive ćelije" je kodiran negdje na nivou elementarnih čestica".
A kako bi drugačije. Svaka ćelija, koja ima DNK, obdarena je svešću, svjesna je sebe i drugih ćelija i u kontaktu je sa Univerzumom, budući da je, zapravo, njegov dio. I iako je broj i raznolikost ćelija u ljudskom tijelu zadivljujući (oko 70 triliona), sve su same sebi slične, kao što su svi procesi koji se odvijaju u ćelijama sami sebi slični. Prema riječima njemačkog naučnika Rolanda Glasera, dizajn bioloških ćelija je "veoma dobro promišljen". Ko je dobro promišljen?
Odgovor je jednostavan: proteini, nukleinske kiseline, žive ćelije i svi biološki sistemi su proizvod kreativne aktivnosti intelektualnog Kreatora.
Ono što je zanimljivo: na atomskom nivou ne postoje razlike između hemijskog sastava organskog i neorganskog sveta. Drugim riječima, na nivou atoma, ćelija je stvorena od istih elemenata kao i neživa priroda. Razlike se nalaze na molekularnom nivou. U živim tijelima, uz neorganske tvari i vodu, nalaze se i proteini, ugljikohidrati, masti, nukleinske kiseline, enzim ATP sintaza i druga organska jedinjenja male molekularne težine.
Do danas, ćelija je doslovno rastavljena na atome u svrhu proučavanja. Međutim, nije moguće stvoriti barem jednu živu ćeliju, jer stvoriti ćeliju znači stvoriti česticu živog Univerzuma. Akademik V.P. Kaznacheev smatra da je "ćelija kosmoplanetarni organizam... Ljudske ćelije su određeni sistemi eterično-torzionih biokolajdera. U tim biokolajderima se dešavaju nama nepoznati procesi, materijalizacija kosmičkih oblika tokova, njihova kosmička transformacija i zbog toga čestice se materijalizuju".
Voda.
Skoro 80% ćelijske mase čini voda. Prema doktoru biologije S. Zeninu, voda je zbog svoje klasterske strukture informaciona matrica za upravljanje biohemijskim procesima. Osim toga, voda je primarna "meta" s kojom djeluju oscilacije zvučne frekvencije. Uređenost stanične vode je toliko visoka (blizu uređenosti kristala) da se naziva tečnim kristalom.
Vjeverice.
Proteini igraju važnu ulogu u biološkom životu. Ćelija sadrži nekoliko hiljada proteina koji su jedinstveni za ovu vrstu ćelija (sa izuzetkom matičnih ćelija). Sposobnost sintetiziranja vlastitih proteina nasljeđuje se od ćelije do ćelije i opstaje tokom života. Tokom života ćelije, proteini postepeno menjaju svoju strukturu, njihova funkcija je narušena. Ovi istrošeni proteini se uklanjaju iz ćelije i zamenjuju novima, zahvaljujući čemu je očuvana vitalna aktivnost ćelije.
Prije svega, ističemo građevnu funkciju proteina, jer su oni građevinski materijal koji čini membrane stanica i stanične organele, zidove krvnih žila, tetive, hrskavicu itd.
Signalna funkcija proteina je izuzetno zanimljiva. Ispostavilo se da su proteini u stanju da služe kao signalne supstance, prenoseći signale između tkiva, ćelija ili organizama. Signalnu funkciju obavljaju hormonski proteini. Ćelije mogu komunicirati jedna s drugom na daljinu koristeći signalne proteine koji se prenose kroz međućelijsku supstancu.
Proteini također imaju motoričku funkciju. Sve vrste pokreta za koje su ćelije sposobne, kao što je kontrakcija mišića, izvode se posebnim kontraktilnim proteinima. Proteini također obavljaju transportnu funkciju. Oni su u stanju da vežu različite supstance i prenose ih sa jednog mesta u ćeliji na drugo. Na primjer, krvni protein hemoglobin vezuje kisik i prenosi ga u sva tkiva i organe tijela. Osim toga, proteini imaju i zaštitnu funkciju. Kada se strani proteini ili ćelije unose u tijelo, u njemu se proizvode posebni proteini koji vežu i neutraliziraju strane stanice i tvari. I konačno, energetska funkcija proteina je da se pri potpunom razgradnji 1 g proteina oslobađa energija u količini od 17,6 kJ.
Struktura ćelije.
Ćelija se sastoji od tri neraskidivo povezana dijela: membrane, citoplazme i jezgra, a struktura i funkcija jezgra u različitim periodima života ćelije su različite. Jer život ćelije uključuje dva perioda: diobu, uslijed koje nastaju dvije kćeri ćelije, i period između dioba, koji se naziva interfaza.
Ćelijska membrana je u direktnoj interakciji sa spoljašnjim okruženjem i u interakciji sa susednim ćelijama. Sastoji se od vanjskog sloja i plazma membrane smještene ispod. Površinski sloj životinjskih ćelija naziva se glikokali. Povezuje ćelije sa spoljašnjim okruženjem i sa svim materijama koje ga okružuju. Njegova debljina je manja od 1 mikrona.
Struktura ćelije
Ćelijska membrana je veoma važan dio ćelije. Drži zajedno sve ćelijske komponente i omeđuje spoljašnje i unutrašnje okruženje.
Postoji stalna razmjena tvari između stanica i vanjskog okruženja. Voda, razne soli u obliku pojedinačnih jona, neorganski i organski molekuli ulaze u ćeliju iz spoljašnje sredine. Produkti metabolizma, kao i tvari koje se sintetiziraju u ćeliji: proteini, ugljikohidrati, hormoni, koji se proizvode u stanicama različitih žlijezda, izlučuju se u vanjsko okruženje kroz membranu iz stanice. Transport supstanci je jedna od glavnih funkcija plazma membrane.
Citoplazma- unutrašnji polutečni medij u kojem se odvijaju glavni metabolički procesi. Nedavne studije su pokazale da citoplazma nije neka vrsta rješenja, čije komponente međusobno komuniciraju u nasumičnim sudarima. Može se uporediti sa želeom, koji počinje da "drhti" kao odgovor na spoljne uticaje. Tako citoplazma percipira i prenosi informacije.
U citoplazmi se nalaze jezgro i različite organele koje su njome ujedinjene u jednu cjelinu, što osigurava njihovu interakciju i aktivnost ćelije kao jedinstvenog integralnog sistema. Jezgro se nalazi u centralnom dijelu citoplazme. Cijela unutrašnja zona citoplazme ispunjena je endoplazmatskim retikulumom, koji je ćelijski organoid: sistem tubula, vezikula i "cisterni" omeđenih membranama. Endoplazmatski retikulum je uključen u metaboličke procese, obezbeđujući transport supstanci iz okoline u citoplazmu i između pojedinačnih intracelularnih struktura, ali njegova glavna funkcija je učešće u sintezi proteina, koja se odvija u ribosomima. - mikroskopska tijela okruglog oblika prečnika 15-20 nm. Sintetizirani proteini se prvo akumuliraju u kanalima i šupljinama endoplazmatskog retikuluma, a zatim transportuju do organela i staničnih mjesta gdje se troše.
Osim proteina, citoplazma sadrži i mitohondrije, mala tijela veličine 0,2-7 mikrona, koja se nazivaju "elektrane" stanica. Redoks reakcije se javljaju u mitohondrijima, dajući ćelijama energiju. Broj mitohondrija u jednoj ćeliji varira od nekoliko do nekoliko hiljada.
Core- vitalni dio ćelije, kontroliše sintezu proteina i preko njih sve fiziološke procese u ćeliji. U jezgri ćelije koja se ne dijeli razlikuje se nuklearna membrana, nuklearni sok, nukleolus i hromozomi. Kroz nuklearni omotač dolazi do kontinuirane izmjene tvari između jezgre i citoplazme. Ispod nuklearnog omotača - nuklearni sok (polutečna tvar), koji sadrži nukleolus i hromozome. Nukleolus je gusto zaobljeno tijelo, čije dimenzije mogu varirati u velikoj mjeri, od 1 do 10 mikrona i više. Sastoji se uglavnom od ribonukleoproteina; učestvuje u formiranju ribozoma. Obično se u ćeliji nalaze 1-3 jezgre, ponekad i do nekoliko stotina. Nukleol se sastoji od RNK i proteina.
Sa pojavom ćelije, život je nastao na Zemlji!
Nastavlja se...


